You might also like
- 8.1.4.7 Packet Tracer - Subnetting Scenario 1Document4 pages8.1.4.7 Packet Tracer - Subnetting Scenario 1Duscosi100% (1)
- Pepsico Corporate Fact SheetDocument2 pagesPepsico Corporate Fact SheetSobia MurtazaNo ratings yet
- Murray Et Al. Prey Experience and Vulnerability To PredationDocument13 pagesMurray Et Al. Prey Experience and Vulnerability To PredationDennis MurrayNo ratings yet
- Nelsen Et Al 2014 Latrodectus DefensaDocument8 pagesNelsen Et Al 2014 Latrodectus DefensaGabriel MaugeriNo ratings yet
- Prey Preferences of The Lion (Panthera Leo) : J. Zool., Lond. (2005) 267, 309-322Document14 pagesPrey Preferences of The Lion (Panthera Leo) : J. Zool., Lond. (2005) 267, 309-322murkyNo ratings yet
- A Comparison of Nutrient Regulation Between Solitarious and Gregarious Phases of The Specialist Caterpillar, Spodoptera Exempta (Walker)Document10 pagesA Comparison of Nutrient Regulation Between Solitarious and Gregarious Phases of The Specialist Caterpillar, Spodoptera Exempta (Walker)jocyeoNo ratings yet
- La Evolución de La Mirmecofagia y Sus Correlatos en Ranas VenenosasDocument27 pagesLa Evolución de La Mirmecofagia y Sus Correlatos en Ranas VenenosasBrandon HerreraNo ratings yet
- 2009, Borteiro Et Al Food Habits Broad Snouted CaimanDocument6 pages2009, Borteiro Et Al Food Habits Broad Snouted CaimanJenifer BorgesNo ratings yet
- Ireland Et Al. 2007Document7 pagesIreland Et Al. 2007Dennis MurrayNo ratings yet
- Thariq Will Find His WayDocument13 pagesThariq Will Find His WayMUHAMMAD FITRIANDESNo ratings yet
- An Exception To The Rule Common Vampire Bats Do Not LearnDocument5 pagesAn Exception To The Rule Common Vampire Bats Do Not LearnBlanca Flor Tirado SierraNo ratings yet
- rspb20063479 PDFDocument6 pagesrspb20063479 PDFThays SantosNo ratings yet
- Carnivore Coexistence Facilitated by Spatial and Dietary Partitioning and Fine-Scale Behavioural Avoidance in A Semi-Arid EcosystemDocument16 pagesCarnivore Coexistence Facilitated by Spatial and Dietary Partitioning and Fine-Scale Behavioural Avoidance in A Semi-Arid EcosystemИванАнато́лийФранкоNo ratings yet
- Does Host Plant Quality Affect The Oviposition Decisions of An Omnivore?Document12 pagesDoes Host Plant Quality Affect The Oviposition Decisions of An Omnivore?Genève DeshaiesNo ratings yet
- Carrying Capacity of Large African Predators Predictions and Tests. Biological ConservationDocument11 pagesCarrying Capacity of Large African Predators Predictions and Tests. Biological ConservationLina Vanesa Garzon PenaNo ratings yet
- Hamilton 2001Document4 pagesHamilton 2001Ilkem KumruNo ratings yet
- Grooming EnglishDocument13 pagesGrooming Englishluisa fernanda jimenez gutierrezNo ratings yet
- 2018 DureDocument11 pages2018 DurenatymdrNo ratings yet
- Wasko 2012Document9 pagesWasko 2012NacorNo ratings yet
- Manova 4Document18 pagesManova 4geo_gabe_galan5473No ratings yet
- Explosive Breeding Reduces Egg and Tadpole Cannibalism in The Wood FrogDocument9 pagesExplosive Breeding Reduces Egg and Tadpole Cannibalism in The Wood FrogCarolina Gomez SernaNo ratings yet
- Ferland-Raymond Et Al 210. Chemical Cues and Prey Behavioural PlasticityDocument9 pagesFerland-Raymond Et Al 210. Chemical Cues and Prey Behavioural PlasticityDennis MurrayNo ratings yet
- Swart Taylor 2004 - Belostomatids Tadpole BufoDocument13 pagesSwart Taylor 2004 - Belostomatids Tadpole Bufooséias martins magalhãesNo ratings yet
- Comparative Analysis of The Diversity of Bird Species in Rural and Urban Areas of The Tirunelveli District, Tamil NaduDocument10 pagesComparative Analysis of The Diversity of Bird Species in Rural and Urban Areas of The Tirunelveli District, Tamil Nadujerzyk.w.locieNo ratings yet
- Voss 2013Document6 pagesVoss 2013jose.1711717678No ratings yet
- The Importance of MotivationDocument26 pagesThe Importance of MotivationNovin TeristiandyNo ratings yet
- Claremont Et Al., 2012Document13 pagesClaremont Et Al., 2012Ricardo López AlonsoNo ratings yet
- Pages de Miller 2Document1 pagePages de Miller 2Irak KrozNo ratings yet
- Damselfly Larvae Learn To Recognize Predators From Chemical Cues in The Predator's DietDocument6 pagesDamselfly Larvae Learn To Recognize Predators From Chemical Cues in The Predator's DietIlkem KumruNo ratings yet
- Ambient Temperature Influences Birds Decisions To Eat To 2013 Animal BehaviDocument8 pagesAmbient Temperature Influences Birds Decisions To Eat To 2013 Animal BehaviAnnie van BlaricumNo ratings yet
- Who Dares, Learns: Chemical Inspection Behaviour and Acquired Predator Recognition in A Characin FishDocument7 pagesWho Dares, Learns: Chemical Inspection Behaviour and Acquired Predator Recognition in A Characin FishharisankarhsNo ratings yet
- How Avian Nest Site Selection Responds To Predation Risk Testing An 'Adaptive Peak Hypothesis' - Journal of Animal Ecology - 2012Document12 pagesHow Avian Nest Site Selection Responds To Predation Risk Testing An 'Adaptive Peak Hypothesis' - Journal of Animal Ecology - 2012carlos murciaNo ratings yet
- Extrapair Paternity and Nest Parasitism in Two Sympatric QuailDocument10 pagesExtrapair Paternity and Nest Parasitism in Two Sympatric QuailConrad ChongNo ratings yet
- Eat Your Heart Out: Choice and Handling of Novel Toxic Prey by Predatory Water RatsDocument5 pagesEat Your Heart Out: Choice and Handling of Novel Toxic Prey by Predatory Water RatsRudson RomeroNo ratings yet
- Evolution 2014 Gibson AKDocument28 pagesEvolution 2014 Gibson AKAlfredo PerettiNo ratings yet
- Prey Preference of Large Carnivores in Anamalai Tiger Reserve, IndiaDocument11 pagesPrey Preference of Large Carnivores in Anamalai Tiger Reserve, IndiaSAVAGE BATTLENo ratings yet
- Composition, Abundance and Feeding Guilds of Macroinvertebrates in Lake Kenyatta, KenyaDocument5 pagesComposition, Abundance and Feeding Guilds of Macroinvertebrates in Lake Kenyatta, KenyaAndrea GonzálezNo ratings yet
- Baudouin Ecology of Snakes Paper - December 2023Document8 pagesBaudouin Ecology of Snakes Paper - December 2023benedict.callebautNo ratings yet
- Animal Domestication and Behavior - F Ch13Document6 pagesAnimal Domestication and Behavior - F Ch13FuligemzNo ratings yet
- HamiltonEtAl 2012Document9 pagesHamiltonEtAl 2012Victor Isaac Perez soteloNo ratings yet
- Meta-Analysis of Foraging and Predation RiskDocument8 pagesMeta-Analysis of Foraging and Predation RiskJennifer VerdolinNo ratings yet
- Downes and Shine (2001b)Document11 pagesDownes and Shine (2001b)Ruben FernandezNo ratings yet
- Captivity Effects On Wide-Ranging CarnivoresDocument2 pagesCaptivity Effects On Wide-Ranging CarnivoresMiguel LessaNo ratings yet
- Eco and Evo 2011Document15 pagesEco and Evo 2011MarcoNo ratings yet
- The Role of Chemical Alarm Signals in Facilitating Learned Recognition of Novel Chemical Cues in A Coral Reef FishDocument7 pagesThe Role of Chemical Alarm Signals in Facilitating Learned Recognition of Novel Chemical Cues in A Coral Reef FishMuliari AyiNo ratings yet
- Gatti Et Al - 2006 - Diet of Two Sympatric Carnivores Cerdocyon and ProcyonDocument4 pagesGatti Et Al - 2006 - Diet of Two Sympatric Carnivores Cerdocyon and ProcyonRita De Cassia BianchiNo ratings yet
- Vences A134 PDFDocument12 pagesVences A134 PDFLucìa SolerNo ratings yet
- Eat or Be EatenDocument11 pagesEat or Be EatenMatthew Peter ClareNo ratings yet
- Hamer Et Al 2006 Carrion V FruitDocument9 pagesHamer Et Al 2006 Carrion V FruitKellyta RodriguezNo ratings yet
- Imprinted Species Recognition Lasts For Life in Free-Living Great Tits and Blue TitsDocument7 pagesImprinted Species Recognition Lasts For Life in Free-Living Great Tits and Blue TitsMohammed KouidriNo ratings yet
- Conservation Genetics: Robin AllabyDocument43 pagesConservation Genetics: Robin AllabyHarry DouglasNo ratings yet
- Evaluating Multiple Arthropod Taxa As Indicators of InvertebrateDocument15 pagesEvaluating Multiple Arthropod Taxa As Indicators of InvertebrateCelular DanielaNo ratings yet
- Roussel Kozon M2eftDocument9 pagesRoussel Kozon M2eftdaniel naoeNo ratings yet
- Lillywhite & Brischoux, 2011Document6 pagesLillywhite & Brischoux, 2011Beatriz BritoNo ratings yet
- Comportamientos AntidepredadorDocument6 pagesComportamientos AntidepredadorMaria NaudeauNo ratings yet
- Behavioural Response To Prey Availability: Population ResilienceDocument1 pageBehavioural Response To Prey Availability: Population Resiliencemamillapalli sri harshaNo ratings yet
- An Integrated Comparison of Captive-Bred and Wild Atlantic Salmon (Salmo Salar) : Implications For Supportive Breeding ProgramsDocument11 pagesAn Integrated Comparison of Captive-Bred and Wild Atlantic Salmon (Salmo Salar) : Implications For Supportive Breeding ProgramsalchemyengrNo ratings yet
- WillifeDocument22 pagesWillifeMaría José de la BárcenaNo ratings yet
- Apex Predators - Arian Wallach, William Ripple Et Al.Document9 pagesApex Predators - Arian Wallach, William Ripple Et Al.Taw GeimNo ratings yet
- Animal Behaviour: Anne Hilborn, Nathalie Pettorelli, C. David L. Orme, Sarah M. DurantDocument6 pagesAnimal Behaviour: Anne Hilborn, Nathalie Pettorelli, C. David L. Orme, Sarah M. DurantMiguel SalNo ratings yet
- Fraser Et Al 2008Document10 pagesFraser Et Al 2008kevin coquitoNo ratings yet
- Merlin Legend 10bsDocument189 pagesMerlin Legend 10bsDuscosiNo ratings yet
- Antitank MineengDocument4 pagesAntitank MineengDuscosiNo ratings yet
- Export Ambitions MECAR Develops New Ammunition RangeDocument8 pagesExport Ambitions MECAR Develops New Ammunition RangeDuscosi100% (1)
- It1 2014-15 - Unit 1 - Accommodation - 1 R-W-L-P-SDocument44 pagesIt1 2014-15 - Unit 1 - Accommodation - 1 R-W-L-P-SPaco Melero RomeroNo ratings yet
- Marketing Plan of Pizza HutDocument11 pagesMarketing Plan of Pizza HutCindy LakhramNo ratings yet
- Action PlanDocument2 pagesAction PlanNadosh HijaziNo ratings yet
- GCS 322 Colores TexturasDocument24 pagesGCS 322 Colores TexturasPrueba PruebaNo ratings yet
- Day Date Time Location Country: Estimation For MealDocument4 pagesDay Date Time Location Country: Estimation For MealSharifah NorsukhairinNo ratings yet
- English 1Document3 pagesEnglish 1crest810No ratings yet
- Outcomes UpperInt VocabBuilder Unit7 0Document7 pagesOutcomes UpperInt VocabBuilder Unit7 0ronaldobatista_engNo ratings yet
- The Dice FramesDocument114 pagesThe Dice FramesValentina KikerkovaNo ratings yet
- New Microsoft Office PowerPoint PresentationDocument27 pagesNew Microsoft Office PowerPoint PresentationGaini ChakravarthyNo ratings yet
- Cernovich, Mike - Snow Crash - Virus, Drug, or ReligionDocument3 pagesCernovich, Mike - Snow Crash - Virus, Drug, or ReligionPeter HeftNo ratings yet
- Karnataka VAT RatesDocument28 pagesKarnataka VAT RatesDeepak Ravindran0% (1)
- Revision Spectrum 3 - Unit 3Document2 pagesRevision Spectrum 3 - Unit 3Cati Lara Cazorla88% (8)
- Coffee Industry (IB Assignment)Document6 pagesCoffee Industry (IB Assignment)Aayush RastogiNo ratings yet
- SUGAR FACTORY and EnvironmentDocument35 pagesSUGAR FACTORY and EnvironmentSudeep singhNo ratings yet
- The HS Classification of Plant-Based Meat - Border BeeDocument7 pagesThe HS Classification of Plant-Based Meat - Border BeeforuzzNo ratings yet
- RaypeatFourm StuffDocument243 pagesRaypeatFourm StuffJoekkk100% (1)
- BOC List of Importers For Suspension-19-January 2017Document9 pagesBOC List of Importers For Suspension-19-January 2017PortCalls100% (1)
- GRADE 11-Health Fitness Wellness Powerpoint 2014Document109 pagesGRADE 11-Health Fitness Wellness Powerpoint 2014Jessieann Balmaceda CabanganNo ratings yet
- Small BusinessDocument18 pagesSmall BusinessWasiur RahmanNo ratings yet
- Answer Key Bar ManagementDocument6 pagesAnswer Key Bar ManagementEmerson CruzNo ratings yet
- Explanation Text How To Make Chocolate: Grestia Angraini Panggabean XA9 Sman 1 Matauli PandanDocument15 pagesExplanation Text How To Make Chocolate: Grestia Angraini Panggabean XA9 Sman 1 Matauli PandanGrestiaangraini PanggabeanNo ratings yet
- Microsoft Word - Vegetables Phonetic Transcription Photos CrosswordDocument35 pagesMicrosoft Word - Vegetables Phonetic Transcription Photos CrosswordCarlosaradasNo ratings yet
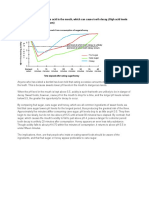
- Eating Sweet Foods Produces Acid in The Mouth, Which Can Cause Tooth Decay. (High Acid Levels Are Measured by Low PH Values)Document19 pagesEating Sweet Foods Produces Acid in The Mouth, Which Can Cause Tooth Decay. (High Acid Levels Are Measured by Low PH Values)Munawir Rahman IslamiNo ratings yet
- Teambuilding Day Proposal Group1Document5 pagesTeambuilding Day Proposal Group1api-580021534No ratings yet
- Costa Cálida Chronicle February 2016Document92 pagesCosta Cálida Chronicle February 2016costacalidachronicleNo ratings yet
- Msds Uq160 enDocument5 pagesMsds Uq160 enAnnalisa GiammòNo ratings yet
- Business Research Project - Piyush's ProjectDocument38 pagesBusiness Research Project - Piyush's ProjectPrerna Goel0% (1)
- JALIL - FLATWARESWritten - Report Topic 9Document2 pagesJALIL - FLATWARESWritten - Report Topic 9Mark Lester TanguanNo ratings yet
- Toxicogenic Insects by MIAN NASiR MEHMOODDocument15 pagesToxicogenic Insects by MIAN NASiR MEHMOODMIAN NASIR100% (2)