You might also like
- OD126164812918973000Document1 pageOD126164812918973000Manoj SirsatNo ratings yet
- Geo WorkbookDocument2 pagesGeo WorkbookKyi Phyu ThantNo ratings yet
- DIET Hassan Science EM Q Bank 2022-23Document60 pagesDIET Hassan Science EM Q Bank 2022-23masoodmohsinNo ratings yet
- Determination of G-Values For Use in SMOGG Gas Generation CalculationsDocument77 pagesDetermination of G-Values For Use in SMOGG Gas Generation CalculationsGaël HenriotNo ratings yet
- 24-Port 10/100/1000Base-T Layer 2+ Full Management Hi T + 4-Port 10G SFP+ High Power Poe SwitchDocument4 pages24-Port 10/100/1000Base-T Layer 2+ Full Management Hi T + 4-Port 10G SFP+ High Power Poe Switchro yenNo ratings yet
- Kizilayin Modernlesme Sahnesinden TasralDocument176 pagesKizilayin Modernlesme Sahnesinden TasralFiseqs MediaNo ratings yet
- PCWB-2316 FLXL Black N100-IdDocument4 pagesPCWB-2316 FLXL Black N100-IdFindora InternusaNo ratings yet
- Concept of State and Regional FinanceDocument15 pagesConcept of State and Regional FinancePutri SariNo ratings yet
- Sda A4 PrintDocument29 pagesSda A4 PrintManoj JariwalaNo ratings yet
- VESDA Sensepoint XCL Large Bore Installation Manual A4Document50 pagesVESDA Sensepoint XCL Large Bore Installation Manual A4Anonymous Mw2nwE6AjNo ratings yet
- Top Fashion Designing Institutes in BhubaneswarDocument3 pagesTop Fashion Designing Institutes in BhubaneswarDigitalNo ratings yet
- 1896-Article Text-7540-1-10-20220308Document13 pages1896-Article Text-7540-1-10-20220308International Health ReviewNo ratings yet
- Pod HandlerDocument13 pagesPod HandlerASWINNo ratings yet
- Cambridge International AS & A Level: CHEMISTRY 9701/22Document16 pagesCambridge International AS & A Level: CHEMISTRY 9701/22The ImmortalNo ratings yet
- 50 de ThiDocument230 pages50 de ThiMinh TâmNo ratings yet
- Boys Tennis UCT 2023 MergedDocument11 pagesBoys Tennis UCT 2023 MergedChris NNo ratings yet
- LAT323 Extra Credit 5Document6 pagesLAT323 Extra Credit 5jam skiNo ratings yet
- General Luna - QuezonDocument1 pageGeneral Luna - QuezonroseannurakNo ratings yet
- (765 KV, 400kV and 220 KV - Existing & Proposed Network) As On 31.3.2021Document1 page(765 KV, 400kV and 220 KV - Existing & Proposed Network) As On 31.3.2021Lohith BBNo ratings yet
- Sanchez Et Al 2010Document12 pagesSanchez Et Al 2010minervaNo ratings yet
- FCE Writing 15 - ArticleDocument4 pagesFCE Writing 15 - Articlepingu70No ratings yet
- 9 English Eng PP 2023 24 1Document12 pages9 English Eng PP 2023 24 1rahulbispur1No ratings yet
- Exhibit A UN Ration Scale 2018 Jan2019Document10 pagesExhibit A UN Ration Scale 2018 Jan2019Gagan Sai ReddyNo ratings yet
- SB Roadmap B1 1Document161 pagesSB Roadmap B1 1Carmen Flores AloyNo ratings yet
- 1 s2.0 S2352396416304108 MainDocument11 pages1 s2.0 S2352396416304108 MainLindia PrabhaswariNo ratings yet
- Matrix Gas Flow Through Impermeable Rocks-Shales and Tight Sandstone PDFDocument28 pagesMatrix Gas Flow Through Impermeable Rocks-Shales and Tight Sandstone PDFẞálôûçh SãmâdNo ratings yet
- Local Table - May 2023Document6 pagesLocal Table - May 2023inforumdocsNo ratings yet
- Sim Automotive-Industries 1899-12 1 4Document17 pagesSim Automotive-Industries 1899-12 1 4Jonathan BachmannNo ratings yet
- 25 InabsentiaDocument272 pages25 InabsentiaANAND MLNo ratings yet
- Https Onetimeregn - Haryana.gov - in PrintApp - AspxDocument3 pagesHttps Onetimeregn - Haryana.gov - in PrintApp - AspxYogita TanwarNo ratings yet
- Kims HospitalDocument547 pagesKims Hospitalmanish.wbsNo ratings yet
- Nylon Carpet. Harrington Catalogue.2022Document41 pagesNylon Carpet. Harrington Catalogue.2022Nikhil Chawla100% (1)
- 2022 05 11 Russian ADR+GDRsDocument1 page2022 05 11 Russian ADR+GDRsSouthey CapitalNo ratings yet
- NM - Vectors and Scalars - Lesson ADocument34 pagesNM - Vectors and Scalars - Lesson Ajohn jkillerzsNo ratings yet
- Sfantul Ignatie Briancianinov Despre Vedenii Duhuri Si MinuniDocument88 pagesSfantul Ignatie Briancianinov Despre Vedenii Duhuri Si MinunigeorgetacaprarescuNo ratings yet
- The LNM Institute of Information Technology, Jaipur: Computer Networks Lab Lab Assignment 1 SolutionDocument181 pagesThe LNM Institute of Information Technology, Jaipur: Computer Networks Lab Lab Assignment 1 SolutionHitesh GoyalNo ratings yet
- 55 Perintah Dasar MikroTik Paling LengkapDocument13 pages55 Perintah Dasar MikroTik Paling Lengkapberry zhubairNo ratings yet
- Imaging in Osteoarthritis 2021 YjocaDocument22 pagesImaging in Osteoarthritis 2021 Yjocaal malikNo ratings yet
- Niva PB ReAssure SS v5Document2 pagesNiva PB ReAssure SS v5samdsozaNo ratings yet
- JLL Business Parks - Rising To The Challenge 2014Document24 pagesJLL Business Parks - Rising To The Challenge 2014Kolehiyo Ng Guiguinto JpiaNo ratings yet
- EuropaDocument5 pagesEuropageorge paunNo ratings yet
- PhonicsDocument31 pagesPhonicsmh0% (1)
- Unit-04 5Document28 pagesUnit-04 5Balaram PaulNo ratings yet
- Solved ISRO Scientist or Engineer Refrigeration & Air-Conditioning 2013 Paper With SolutionsDocument22 pagesSolved ISRO Scientist or Engineer Refrigeration & Air-Conditioning 2013 Paper With SolutionsD Eswar PrasadNo ratings yet
- International Trade, Finance & Investment Report - Guidelines Deadline: 21 January 2022Document7 pagesInternational Trade, Finance & Investment Report - Guidelines Deadline: 21 January 2022haseeb ahmedNo ratings yet
- 11 AppendixDocument19 pages11 AppendixГеорги ПренчевNo ratings yet
- United States Patent: (10) Patent No .: US 10, 278, 244 B1Document45 pagesUnited States Patent: (10) Patent No .: US 10, 278, 244 B1Roberto AbregoNo ratings yet
- 11-2022-23-02-001 (2) Agra BillDocument1 page11-2022-23-02-001 (2) Agra BillMeenakshi VermaNo ratings yet
- Menu 259Document2 pagesMenu 259DanielaSalinasNo ratings yet
- Falaknuma Exp Third Ac (3A)Document2 pagesFalaknuma Exp Third Ac (3A)Abhaya Kumar NayakNo ratings yet
- A1 Customer Information FormDocument2 pagesA1 Customer Information FormJimmy TangonanNo ratings yet
- Round Robin ReadingDocument24 pagesRound Robin ReadingBriones JRNo ratings yet
- Starch Based Biodegradable Blends - A ReviewDocument18 pagesStarch Based Biodegradable Blends - A Reviewsoheil farshbafNo ratings yet
- 3 Is Learning Information Sheet in Technical Drafting Third Week 1Document11 pages3 Is Learning Information Sheet in Technical Drafting Third Week 1Cherie BalentozaNo ratings yet
- Di Ethylene Glycol - RIL - MSDSDocument10 pagesDi Ethylene Glycol - RIL - MSDSKannan NainarNo ratings yet
- Orca Share Media1672898478819 7016644781308336021Document4 pagesOrca Share Media1672898478819 7016644781308336021Vanessa Karikitan EstorNo ratings yet
- Check List For Immigration Visa NewDocument1 pageCheck List For Immigration Visa NewХарш Навнітбхаі НімаватNo ratings yet
- A Brief Study On Structural Use & High Performance of Jute in Different Applications and Its Prospects in Future UsesDocument66 pagesA Brief Study On Structural Use & High Performance of Jute in Different Applications and Its Prospects in Future UsesMosharaf HossainNo ratings yet
- What Is Molecular FarmingDocument20 pagesWhat Is Molecular Farmingmcops20110% (1)
- Exploitation of Ant TechniquesDocument12 pagesExploitation of Ant TechniquessudinNo ratings yet
- Bakar CD RobiwijayaDocument60 pagesBakar CD RobiwijayaAsep Ribbat MNo ratings yet
- Personal StatementDocument1 pagePersonal StatementTaiwo Oyeniyi100% (2)
- Chapter 7 Africa South of The SaharaDocument40 pagesChapter 7 Africa South of The Saharafuck ypouNo ratings yet
- Effect of Pre-Cooling Injection Site On Pain Perception in Pediatric Dentistry: "A Randomized Clinical Trial"Document6 pagesEffect of Pre-Cooling Injection Site On Pain Perception in Pediatric Dentistry: "A Randomized Clinical Trial"Ummi AzizahNo ratings yet
- Jpog August 2013 IdDocument51 pagesJpog August 2013 IdHasyim PurwadiNo ratings yet
- Management of DiabetesDocument164 pagesManagement of Diabetesvivek_win95No ratings yet
- SunCoBe ShampooDocument24 pagesSunCoBe ShampooDong YulNo ratings yet
- GHH Horoscope PDFDocument46 pagesGHH Horoscope PDFRakesh Kumar BeheraNo ratings yet
- 1 s2.0 S0828282X22001271 MainDocument11 pages1 s2.0 S0828282X22001271 MainLilianne Mbengani LaranjeiraNo ratings yet
- Plasmodium Infection (Malaria) : Medical ParasitologyDocument7 pagesPlasmodium Infection (Malaria) : Medical ParasitologyBalisi Manuel FranciscoNo ratings yet
- (1913) List of Licensed EmbalmersDocument96 pages(1913) List of Licensed EmbalmersHerbert Hillary Booker 2nd0% (1)
- A Study On Knowledge and Practice of Mothers of Under-Five ChildrenDocument6 pagesA Study On Knowledge and Practice of Mothers of Under-Five ChildrenRirin PurbaNo ratings yet
- HIV in South AfricaDocument17 pagesHIV in South AfricaMadelineNo ratings yet
- Pyelonephritis: Dr. Shraddha Koirala Department of Pathology NMCTHDocument26 pagesPyelonephritis: Dr. Shraddha Koirala Department of Pathology NMCTHUrusha MagarNo ratings yet
- Anatomy and Physiology of EyelidDocument34 pagesAnatomy and Physiology of Eyelidsushma shrestha100% (1)
- Nosebleed and PoisoningDocument28 pagesNosebleed and PoisoningRenz100% (2)
- Diet & Nutrition Advanced Ver 1Document52 pagesDiet & Nutrition Advanced Ver 1Ina100% (1)
- The Role of Chronic PainDocument23 pagesThe Role of Chronic PainNovia RambakNo ratings yet
- Announcement Sinas IDAI - Draft 07Document5 pagesAnnouncement Sinas IDAI - Draft 07Hendra JayaNo ratings yet
- Challenges Faced by Families of Autistic Children-153Document5 pagesChallenges Faced by Families of Autistic Children-153Zahoor Ahmad100% (1)
- Dermatology - James G. Marks, Jeffrey J. Miller - Lookingbill and Marks' Principles of Dermatology (2018, Elsevier) (Dragged)Document9 pagesDermatology - James G. Marks, Jeffrey J. Miller - Lookingbill and Marks' Principles of Dermatology (2018, Elsevier) (Dragged)nh2411No ratings yet
- Anesthetic Management of Non-Fasted Patients Requiring Emergency SurgeryDocument17 pagesAnesthetic Management of Non-Fasted Patients Requiring Emergency SurgeryBhi-An BatobalonosNo ratings yet
- CHAPTER 4: Modifiable Risk Factors of Lifestyle Diseases: SmokingDocument4 pagesCHAPTER 4: Modifiable Risk Factors of Lifestyle Diseases: SmokingShimah100% (2)
- 2020 Article 773Document10 pages2020 Article 773bagas umam alwiNo ratings yet
- BCAAs and Body Composition - Art de Vany On LineDocument2 pagesBCAAs and Body Composition - Art de Vany On LineDellNo ratings yet
- Santosh DevDocument45 pagesSantosh Devdevdsantosh100% (1)
- Lecture 5 - Facial Pain and TMJ DiseaseDocument6 pagesLecture 5 - Facial Pain and TMJ DiseaseJeff ChadwickNo ratings yet
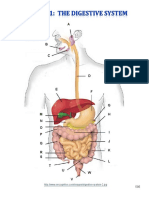
- Digestive System Charts 1Document14 pagesDigestive System Charts 1api-306762527No ratings yet
- Market Potential For Probiotic Nutritional Supplem PDFDocument7 pagesMarket Potential For Probiotic Nutritional Supplem PDFshanmuga8801No ratings yet