You might also like
- American Indian Symbols DictionaryDocument11 pagesAmerican Indian Symbols DictionaryMarjelyn Lavarrete MartirezNo ratings yet
- d100 Side Quest Hooks GM BinderDocument5 pagesd100 Side Quest Hooks GM Bindermiguelanderson190No ratings yet
- Symbiosis A Relationship Where Two Organisms (Of Different Species) Live TogetherDocument5 pagesSymbiosis A Relationship Where Two Organisms (Of Different Species) Live TogetherJr MateoNo ratings yet
- The Origin and Evolution of BirdsDocument20 pagesThe Origin and Evolution of BirdsEmanuelleNo ratings yet
- How To Start A Small Scale Poultry Farm PDFDocument13 pagesHow To Start A Small Scale Poultry Farm PDFnebiyu samuelNo ratings yet
- Discoveries of New Mammal SpeciesDocument7 pagesDiscoveries of New Mammal Species--No ratings yet
- In Harm's Way Dragons! Breeds of The OrientDocument21 pagesIn Harm's Way Dragons! Breeds of The OrientLauraHenson100% (2)
- Insectsin Forensic InvestigationsDocument18 pagesInsectsin Forensic InvestigationsJudicaëlNo ratings yet
- 1 s2.0 S0003347202930174 MainDocument10 pages1 s2.0 S0003347202930174 Mainandreea.dumitrescuNo ratings yet
- Magazine: Microbiology of DeathDocument3 pagesMagazine: Microbiology of DeathElva PatabangNo ratings yet
- ConvergenceDocument10 pagesConvergenceKiss LeventeNo ratings yet
- Dung Beetles Key To Healthy PastureDocument31 pagesDung Beetles Key To Healthy PastureLEIDY VIVIANA GARCIA HERRERANo ratings yet
- Interspecific Interactions Between Adult Aedes Albopictus and Culex Quinquefasciatus (Diptera: Culicidae)Document4 pagesInterspecific Interactions Between Adult Aedes Albopictus and Culex Quinquefasciatus (Diptera: Culicidae)ALEX CHAVIER SILVANo ratings yet
- Research Papers On Birds DiversityDocument6 pagesResearch Papers On Birds Diversityafnkjdhxlewftq100% (1)
- Proposal To Sequence The First Reptilian Genome: The Green Anole Lizard, Anolis CarolinensisDocument21 pagesProposal To Sequence The First Reptilian Genome: The Green Anole Lizard, Anolis CarolinensisJamesComeyJustaBitchNo ratings yet
- Tropicalbiomedicine 2019Document10 pagesTropicalbiomedicine 2019prashantaraj1020No ratings yet
- 0102 0935 Abmvz 69 05 01172 PDFDocument9 pages0102 0935 Abmvz 69 05 01172 PDFLucas VerniereNo ratings yet
- Economic Anth FinalDocument10 pagesEconomic Anth Finalapi-549334775No ratings yet
- The Importance of MotivationDocument26 pagesThe Importance of MotivationNovin TeristiandyNo ratings yet
- García Hernandez Machado 2017 Effectiveness of Maternal Egg Attendance in An Exclusively HarvestmanDocument8 pagesGarcía Hernandez Machado 2017 Effectiveness of Maternal Egg Attendance in An Exclusively Harvestmanlola zavaliaNo ratings yet
- Biological ConservationDocument9 pagesBiological ConservationEfendi SamaleleNo ratings yet
- CockroachesDocument11 pagesCockroachesZahra 9oNo ratings yet
- Stri Wikelski Kays Kasdin Et Al 2007Document6 pagesStri Wikelski Kays Kasdin Et Al 2007Bereket LemmaNo ratings yet
- Landfill Use by Bald Eagles in The Chesapeake Bay RegionDocument12 pagesLandfill Use by Bald Eagles in The Chesapeake Bay RegionImamFadilaNo ratings yet
- Systematics and PhylogeneticsDocument4 pagesSystematics and PhylogeneticslazygemNo ratings yet
- Chapter 2Document7 pagesChapter 2Caryl Mae BocadoNo ratings yet
- Ticks and Tick-Borne DiseasesDocument8 pagesTicks and Tick-Borne DiseasesLørïïs SïerraNo ratings yet
- Alcock 1982 PDFDocument17 pagesAlcock 1982 PDFheimat66No ratings yet
- Poultry Welfare PrakashDocument3 pagesPoultry Welfare PrakashSuresh NepaliNo ratings yet
- Jones and Chapman 2020. The Ethics and Welfare Implications of Keeping Western European Hedgehogs Erinaceus Europaeus in CaptivityDocument18 pagesJones and Chapman 2020. The Ethics and Welfare Implications of Keeping Western European Hedgehogs Erinaceus Europaeus in CaptivityLuciano VerzolaNo ratings yet
- Ecological Importance of Birds - M A Tabur 2010Document6 pagesEcological Importance of Birds - M A Tabur 2010crew90No ratings yet
- Ireland Et Al. 2007Document7 pagesIreland Et Al. 2007Dennis MurrayNo ratings yet
- Venus Flytrap Rarely Traps Its PollinatorsDocument9 pagesVenus Flytrap Rarely Traps Its PollinatorsVivi TorresNo ratings yet
- 2011 Dropping DeadDocument15 pages2011 Dropping Deadaqueeb javeedNo ratings yet
- Wildt, 1997Document11 pagesWildt, 1997Adauto AlvesNo ratings yet
- Bartlow 2016Document7 pagesBartlow 2016Jevinur EffendyNo ratings yet
- Comparative GenomicsDocument6 pagesComparative Genomicshar2dikNo ratings yet
- Chapter 1: Carnivores in Human-Influenced LandscapesDocument1 pageChapter 1: Carnivores in Human-Influenced Landscapesmamillapalli sri harshaNo ratings yet
- The Evolution of Crocodilian Nesting Ecology and Behavior: Christopher M. Murray - Brian I. Crother - J. Sean DoodyDocument19 pagesThe Evolution of Crocodilian Nesting Ecology and Behavior: Christopher M. Murray - Brian I. Crother - J. Sean DoodyFlaviana Tamung02No ratings yet
- Shah Et Al 2003 (Sociality in Lizards) BehaviourDocument14 pagesShah Et Al 2003 (Sociality in Lizards) BehaviourjavieromaNo ratings yet
- Theory Culture Society 2016 Van Dooren 29 52Document24 pagesTheory Culture Society 2016 Van Dooren 29 52Julia HartlineNo ratings yet
- The Definition of SpeciesDocument3 pagesThe Definition of Speciesali7167No ratings yet
- Systematics and PhylogenyDocument5 pagesSystematics and PhylogenyBryan FabroNo ratings yet
- Eat Your Heart Out: Choice and Handling of Novel Toxic Prey by Predatory Water RatsDocument5 pagesEat Your Heart Out: Choice and Handling of Novel Toxic Prey by Predatory Water RatsRudson RomeroNo ratings yet
- Possible Variations To Avoid The Extinction of Dodo (Raphus Cucullatus)Document3 pagesPossible Variations To Avoid The Extinction of Dodo (Raphus Cucullatus)marizNo ratings yet
- Tomberlin Et Al 2017Document18 pagesTomberlin Et Al 2017Jose Alejandro Hernandez BenitezNo ratings yet
- Pagel 2009 - Natural Selection 150 Years OnDocument4 pagesPagel 2009 - Natural Selection 150 Years OnCamilo GarcíaNo ratings yet
- HSC Biology Blue Print of LifeDocument64 pagesHSC Biology Blue Print of LifeJeffrey Avacado LiuNo ratings yet
- Vitt Et Al. 2003 - (History and The Global Ecology of Squamate Reptiles)Document17 pagesVitt Et Al. 2003 - (History and The Global Ecology of Squamate Reptiles)Raissa SiqueiraNo ratings yet
- Carnivore Coexistence Facilitated by Spatial and Dietary Partitioning and Fine-Scale Behavioural Avoidance in A Semi-Arid EcosystemDocument16 pagesCarnivore Coexistence Facilitated by Spatial and Dietary Partitioning and Fine-Scale Behavioural Avoidance in A Semi-Arid EcosystemИванАнато́лийФранкоNo ratings yet
- 49 Insect Foraging in Captive Owl Monkeys Wolovich Et Al 2009Document10 pages49 Insect Foraging in Captive Owl Monkeys Wolovich Et Al 2009Vicky Del PradoNo ratings yet
- WAFT Reading June2013Document2 pagesWAFT Reading June2013lsxmiki9No ratings yet
- Fajfer 2012Document22 pagesFajfer 2012Monika FajferNo ratings yet
- Almiron Et Al 2011 A Failed Swallowing Attempt Upon A Crab-Eating FoxDocument4 pagesAlmiron Et Al 2011 A Failed Swallowing Attempt Upon A Crab-Eating FoxEverton MirandaNo ratings yet
- Eat Your Heart Out - Choice and Handling of Novel Toxic Prey by Predatory Water RatsDocument6 pagesEat Your Heart Out - Choice and Handling of Novel Toxic Prey by Predatory Water RatsJean Carlos MirandaNo ratings yet
- Viperidae (Crotalinae) - A Family and Its NicheDocument6 pagesViperidae (Crotalinae) - A Family and Its NicheCelia RodriguesNo ratings yet
- Animal Welfare Aspects of Outdoor Runs For Laying Hens: A ReviewDocument13 pagesAnimal Welfare Aspects of Outdoor Runs For Laying Hens: A ReviewAlejandra Gomez RuizNo ratings yet
- Fas DietDocument8 pagesFas DietDuscosiNo ratings yet
- Grooming EnglishDocument13 pagesGrooming Englishluisa fernanda jimenez gutierrezNo ratings yet
- Systematics and Evolution of Spiders (Araneae) : Jonathan A. CoddingtonDocument28 pagesSystematics and Evolution of Spiders (Araneae) : Jonathan A. CoddingtonHernanFigueredoNo ratings yet
- Apex Predators - Arian Wallach, William Ripple Et Al.Document9 pagesApex Predators - Arian Wallach, William Ripple Et Al.Taw GeimNo ratings yet
- Genome Resource BanksDocument20 pagesGenome Resource BanksFernanda GouveaNo ratings yet
- Centipedes Vs SalamandersDocument8 pagesCentipedes Vs SalamandersjavieromaNo ratings yet
- ATQ (Bio Lab 12) E7Document3 pagesATQ (Bio Lab 12) E7fizzypurplesodaNo ratings yet
- Barred Owl Field Study - Final Project 2Document6 pagesBarred Owl Field Study - Final Project 2api-486211746No ratings yet
- Final Group Paper 3Document10 pagesFinal Group Paper 3api-486211746No ratings yet
- Final Project OcavalDocument6 pagesFinal Project Ocavalapi-486211746No ratings yet
- Soils Photo ProjectDocument9 pagesSoils Photo Projectapi-486211746No ratings yet
- Esrm 350 PresentationDocument37 pagesEsrm 350 Presentationapi-486211746No ratings yet
- ECA1 - Tests - Language Test 7C - New2018Document6 pagesECA1 - Tests - Language Test 7C - New2018ejakubowNo ratings yet
- EVS Revision Paper Annual Exam 2022Document8 pagesEVS Revision Paper Annual Exam 2022Sachin PatelNo ratings yet
- Soal Farm Animal Kelas 5Document3 pagesSoal Farm Animal Kelas 5Ir SoeparnoNo ratings yet
- ANIMALS - CAMBRIGE VOCABULARY-MOVERS 5 y 6 Audio-1.ppsxDocument31 pagesANIMALS - CAMBRIGE VOCABULARY-MOVERS 5 y 6 Audio-1.ppsxManuel Bustamante CernaNo ratings yet
- Activity Plan 2 Year Old May 11Document6 pagesActivity Plan 2 Year Old May 11Dante GordonNo ratings yet
- Ace Reasoning New AddaDocument359 pagesAce Reasoning New Addajuinarka24112002No ratings yet
- Cimetrols Rfu LabelDocument1 pageCimetrols Rfu Labelwkjg7w7j2fNo ratings yet
- Chicken Anatomy and Physiology: Respiratory System: by Jacquie Jacob and Tony PescatoreDocument4 pagesChicken Anatomy and Physiology: Respiratory System: by Jacquie Jacob and Tony Pescatoreసాయిమహేష్రెడ్డిఆవులNo ratings yet
- Manual of Nearctic Diptera V1 McAlpine Et Al 1981 9 MorphologyDocument80 pagesManual of Nearctic Diptera V1 McAlpine Et Al 1981 9 MorphologyChristophe AvonNo ratings yet
- the-life-of-the-european-bee-eater-đã chuyển đổiDocument3 pagesthe-life-of-the-european-bee-eater-đã chuyển đổiLinh TrangNo ratings yet
- The Cunning Fox and The Clever StorkDocument1 pageThe Cunning Fox and The Clever StorksridharNo ratings yet
- An Assignment On Respiratory System and Sound SystemDocument17 pagesAn Assignment On Respiratory System and Sound SystemMD Taufique HussainNo ratings yet
- Buffalo MosquitoDocument9 pagesBuffalo MosquitoMeldon MacTavishNo ratings yet
- Detailed Lesson Plan 2Document8 pagesDetailed Lesson Plan 2Christine Vallente ArcabalNo ratings yet
- PH-09 (KD 3.9) Proverb & Riddle (PG40) GFormDocument6 pagesPH-09 (KD 3.9) Proverb & Riddle (PG40) GFormLahita AzizahNo ratings yet
- TEENS-5 - Practice Test 2020Document8 pagesTEENS-5 - Practice Test 2020Sofia DuvaNo ratings yet
- Maths Workbook - Basic 1 - 2nd TermDocument66 pagesMaths Workbook - Basic 1 - 2nd TermLateef OlamideNo ratings yet
- Toyota Forklift 8fbm20t To 8fbm35t 8fbmk25t 8fbmk30t Repair ManualDocument23 pagesToyota Forklift 8fbm20t To 8fbm35t 8fbmk25t 8fbmk30t Repair Manualalexhughes210188dzo100% (116)
- ODI 4 Test Unit 5-6Document2 pagesODI 4 Test Unit 5-6SilvanaNo ratings yet
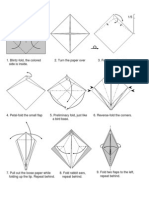
- Dragon OrigamiDocument2 pagesDragon Origamishin_chanNo ratings yet
- Marked 80 Mcqs Poultry BidarDocument11 pagesMarked 80 Mcqs Poultry BidarYasawal WorkingNo ratings yet
- Lighting Program For Small Roaster FlocksDocument3 pagesLighting Program For Small Roaster FlocksAgustino WaitihacheNo ratings yet
- Top 10 Flightless Birds You Won't Believe ExistDocument3 pagesTop 10 Flightless Birds You Won't Believe ExistRidwan AdedejiNo ratings yet
- Luyện ĐọcDocument8 pagesLuyện Đọcdinhduyen610No ratings yet
- Collective Noun Farm: Learning ObjectivesDocument6 pagesCollective Noun Farm: Learning ObjectivesHannah DYNo ratings yet