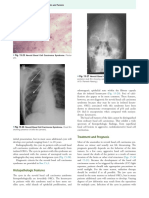
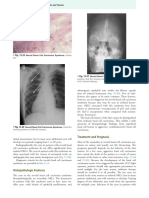
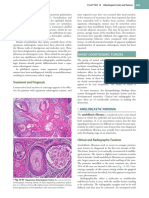
You might also like
- Notes: Heparin-Induced Thrombocytopenia (Hit)Document6 pagesNotes: Heparin-Induced Thrombocytopenia (Hit)MOHIT SHARMANo ratings yet
- Mechanisms of Infectious DiseaseDocument28 pagesMechanisms of Infectious DiseaseHastyar KhalidNo ratings yet
- PeriodontitisDocument32 pagesPeriodontitisDimitrios PapadopoulosNo ratings yet
- Goat Medicine and SurgeryDocument393 pagesGoat Medicine and Surgerydrum75% (4)
- Drug Inspector Previous Exam Paper With Answers PDF APPSCDocument14 pagesDrug Inspector Previous Exam Paper With Answers PDF APPSCSheik Hussain100% (3)
- Yeast Respiration ExperimentDocument13 pagesYeast Respiration ExperimentKayliNo ratings yet
- Cell SignalingDocument53 pagesCell SignalingKaushiki KalravNo ratings yet
- MCQ4FULLDocument139 pagesMCQ4FULLBruta Brutal100% (2)
- Kjim 2016 098Document9 pagesKjim 2016 098Anonymous 7dsX2F8nNo ratings yet
- What Treatment of Tendinopathy: Inflammation or Degeneration?Document4 pagesWhat Treatment of Tendinopathy: Inflammation or Degeneration?Shaun TylerNo ratings yet
- Yu 2016Document21 pagesYu 2016Manuel Esquivel OrtegaNo ratings yet
- Extracellular Vesicles Derived From HumanDocument21 pagesExtracellular Vesicles Derived From HumanMartín MamchurNo ratings yet
- Role of Mast Cells in Periodontal DiseaseDocument26 pagesRole of Mast Cells in Periodontal DiseaseDpartment of PeriodontologyNo ratings yet
- Macrophages in OsteoarthritisDocument8 pagesMacrophages in OsteoarthritisТати ГалаNo ratings yet
- Ledesmamontes 2004Document5 pagesLedesmamontes 2004TORRES POVEDA MARIA XIMENANo ratings yet
- Bioengineering 11 00357Document15 pagesBioengineering 11 00357Shahmeer HassanNo ratings yet
- Cytokine: Mohammad M.Y. Khorasani, Gholamhossein Hassanshahi, Aniela Brodzikowska, Hossein Khorramdelazad TDocument8 pagesCytokine: Mohammad M.Y. Khorasani, Gholamhossein Hassanshahi, Aniela Brodzikowska, Hossein Khorramdelazad TJade LolongNo ratings yet
- Nampath MonositDocument8 pagesNampath MonositEllya Latifah IlyasNo ratings yet
- Artrosis MSCDocument9 pagesArtrosis MSCJob MonobeNo ratings yet
- Reviews: Neutrophil Diversity and Plasticity in Tumour Progression and TherapyDocument19 pagesReviews: Neutrophil Diversity and Plasticity in Tumour Progression and TherapySobek1789No ratings yet
- Artículo 1Document18 pagesArtículo 1AlejandraNo ratings yet
- Zhu 2022Document16 pagesZhu 2022Z MNo ratings yet
- Temporomandibular Joint Pain and Synovial Fluid Analysis: A Review of The LiteratureDocument8 pagesTemporomandibular Joint Pain and Synovial Fluid Analysis: A Review of The Literaturedavidcosmin186832No ratings yet
- Okizaki, 2015Document9 pagesOkizaki, 2015wanda oktariaNo ratings yet
- Journal Kiki - VeldaDocument18 pagesJournal Kiki - VeldaYvonne DoyleNo ratings yet
- Macrophage M1 M2 PolarizationDocument9 pagesMacrophage M1 M2 PolarizationBrilliantNo ratings yet
- Loukov 2018Document9 pagesLoukov 2018Jonathan Billy ChristianNo ratings yet
- Macrophage Mediation in Normal and Diabetic Wound Healing ResponsesDocument17 pagesMacrophage Mediation in Normal and Diabetic Wound Healing ResponsesHarithpriya KannanNo ratings yet
- MMR 9 1 9 PDFDocument7 pagesMMR 9 1 9 PDFIndira Damar PangestuNo ratings yet
- Author's Accepted Manuscript: Experimental Cell ResearchDocument25 pagesAuthor's Accepted Manuscript: Experimental Cell ResearchKarolNo ratings yet
- PEMF - Rheumatoid ArhritisDocument12 pagesPEMF - Rheumatoid ArhritisMáté TóthNo ratings yet
- T Cell Migration in RheumatoidDocument12 pagesT Cell Migration in RheumatoidamurachNo ratings yet
- Inflammation and Its Resolution and The Musculoskeletal SystemDocument16 pagesInflammation and Its Resolution and The Musculoskeletal Systemingrid da silvaNo ratings yet
- Microambiente TumoralDocument2 pagesMicroambiente TumoralesmargarNo ratings yet
- Research Article in Vitro, in VivoDocument20 pagesResearch Article in Vitro, in VivoRachmawati Dian PuspitasariNo ratings yet
- E003586 FullDocument4 pagesE003586 FullawaloeiacidNo ratings yet
- The Effect of Tumor Necrosis Factor (TNF) - To Induce Matrix Metalloproteinase (MMPS) From The Human Dental Pulp, Gingival, and Periodontal Ligament CellsDocument11 pagesThe Effect of Tumor Necrosis Factor (TNF) - To Induce Matrix Metalloproteinase (MMPS) From The Human Dental Pulp, Gingival, and Periodontal Ligament CellsNadya PurwantyNo ratings yet
- Inflmsi JosDocument27 pagesInflmsi Jospramita wahyu dyastiNo ratings yet
- Journal of Periodontology - 2000 - Séguier - Collagen Fibers and Inflammatory Cells in Healthy and Diseased Human GingivalDocument7 pagesJournal of Periodontology - 2000 - Séguier - Collagen Fibers and Inflammatory Cells in Healthy and Diseased Human GingivalLuiz Guilherme FiorinNo ratings yet
- Pharmaceuticals 15 01094 v2Document20 pagesPharmaceuticals 15 01094 v2Faizal Reza PahleviNo ratings yet
- ArtrosisDocument8 pagesArtrosisAdrianNo ratings yet
- Ar FisiopatologiaDocument10 pagesAr FisiopatologiaEdyson SedanoNo ratings yet
- Verde 2Document6 pagesVerde 2Raluca Andreea AxinteNo ratings yet
- McInnes y Schett - 2017 - Pathogenetic Insights From The Treatment of RheumaDocument20 pagesMcInnes y Schett - 2017 - Pathogenetic Insights From The Treatment of Rheumabarrera2001No ratings yet
- Periodontitis: An Inevitable DestructionDocument4 pagesPeriodontitis: An Inevitable DestructionasclepiuspdfsNo ratings yet
- The Cytokine Network Involved in The Host Immune Response To PeriodontitisDocument13 pagesThe Cytokine Network Involved in The Host Immune Response To PeriodontitisGeorgi GugicevNo ratings yet
- Fimmu 13 1003651Document16 pagesFimmu 13 1003651BG BBNo ratings yet
- Immunomodulatory Activities of Periodontal Ligament Stem Cells in Orthodontic Forces-Induced Inflammatory Processes: Current Views and Future PerspectivesDocument8 pagesImmunomodulatory Activities of Periodontal Ligament Stem Cells in Orthodontic Forces-Induced Inflammatory Processes: Current Views and Future Perspectives赵忠琪No ratings yet
- 10 1016@j Biomaterials 2020 120390Document16 pages10 1016@j Biomaterials 2020 120390PuntalusNo ratings yet
- Biomedicines 12 00686Document11 pagesBiomedicines 12 00686jamel-shamsNo ratings yet
- Functional and Phenotypic Changes in Monocytes From Patients With Tuberculosis Are Reversed With TreatmentDocument9 pagesFunctional and Phenotypic Changes in Monocytes From Patients With Tuberculosis Are Reversed With TreatmentCecile E EcheverríaNo ratings yet
- Mitofusin 2 in Macrophages Links Mitochondrial ROS Production, Cytokine Release, Phagocytosis, Autophagy, and Bactericidal ActivityDocument25 pagesMitofusin 2 in Macrophages Links Mitochondrial ROS Production, Cytokine Release, Phagocytosis, Autophagy, and Bactericidal Activityjuan carlos monasterio saezNo ratings yet
- Onionin A Inhibits Ovarian Cancer Progression by Suppressing Cancer Cell Proliferation and The Protumour Function of MacrophagesDocument11 pagesOnionin A Inhibits Ovarian Cancer Progression by Suppressing Cancer Cell Proliferation and The Protumour Function of Macrophageskhangkhang151203No ratings yet
- Potential Anti-Inflammatory Treatments For Chronic Wounds: Mckelvey K, Xue M, Whitmont K, Shen K, Cooper A & Jackson CDocument4 pagesPotential Anti-Inflammatory Treatments For Chronic Wounds: Mckelvey K, Xue M, Whitmont K, Shen K, Cooper A & Jackson CAnonymous IruFyBNo ratings yet
- Cytokine: Mili Gupta, Rashi Chaturvedi, Ashish JainDocument6 pagesCytokine: Mili Gupta, Rashi Chaturvedi, Ashish JainShreyaNo ratings yet
- Magnetotherapy: The Quest For Tendon Regeneration: Tamagno Pesqueira Raquel Costa-Almeida Manuela E. GomesDocument11 pagesMagnetotherapy: The Quest For Tendon Regeneration: Tamagno Pesqueira Raquel Costa-Almeida Manuela E. GomesDayana VILLANUEVA CONTRERASNo ratings yet
- The Role of Inflammation in The Pathogenesis of OsteoarthritisDocument19 pagesThe Role of Inflammation in The Pathogenesis of Osteoarthritiscursotplo2021No ratings yet
- Angio GenesisDocument8 pagesAngio GenesisSamarTharwatNo ratings yet
- 2016 - Observing The Development of The ATM in EmbryonicDocument9 pages2016 - Observing The Development of The ATM in EmbryonicJULIA SANFURGO VILLARROELNo ratings yet
- The Role of Prophylactic Ibuprofen and N-Acetylcysteine On The Level of Cytokines in Periapical Exudates and The Post-Treatment PainDocument6 pagesThe Role of Prophylactic Ibuprofen and N-Acetylcysteine On The Level of Cytokines in Periapical Exudates and The Post-Treatment PainNadya PurwantyNo ratings yet
- Int Jurnal OncoDocument9 pagesInt Jurnal OncoWida MarianeNo ratings yet
- ACVR1R206H Extends Inflammatory Responses in Human Induced Pluripote 2021 BoDocument17 pagesACVR1R206H Extends Inflammatory Responses in Human Induced Pluripote 2021 BodianaNo ratings yet
- Cancers: Role of Inflammation and Oxidative Stress Mediators in GliomasDocument20 pagesCancers: Role of Inflammation and Oxidative Stress Mediators in GliomasVictor HaidarNo ratings yet
- s41420 021 00418 yDocument15 pagess41420 021 00418 yValen EstevezNo ratings yet
- Review: Mesenchymal Stromal Cells: Sensors and Switchers of InflammationDocument11 pagesReview: Mesenchymal Stromal Cells: Sensors and Switchers of InflammationlilisNo ratings yet
- Pemphigus in Cardiac PatientsDocument31 pagesPemphigus in Cardiac Patientszendah123No ratings yet
- Mesothelioma: From Research to Clinical PracticeFrom EverandMesothelioma: From Research to Clinical PracticeGiovanni Luca CeresoliNo ratings yet
- The Problems Preventing Progress in Adhesive Restorative DentistryDocument6 pagesThe Problems Preventing Progress in Adhesive Restorative DentistrygermanNo ratings yet
- Resveratrol Alleviates Temporomandibular Joint Inflammatory Pain by Recovering Disturbed Gut MicrobiotaDocument20 pagesResveratrol Alleviates Temporomandibular Joint Inflammatory Pain by Recovering Disturbed Gut MicrobiotagermanNo ratings yet
- Meta 2.1 Ocampo LibroDocument21 pagesMeta 2.1 Ocampo LibrogermanNo ratings yet
- The Use of Pharmacologic Agents in The Management of Temporomandibular Joint DisorderDocument7 pagesThe Use of Pharmacologic Agents in The Management of Temporomandibular Joint DisordergermanNo ratings yet
- Ribeiro 2020Document8 pagesRibeiro 2020germanNo ratings yet
- Meta 2.1 Ocampo Libro (15-21)Document7 pagesMeta 2.1 Ocampo Libro (15-21)germanNo ratings yet
- Meta 2.1 Ocampo Libro (08-14)Document7 pagesMeta 2.1 Ocampo Libro (08-14)germanNo ratings yet
- Meta 2.1 Ocampo Libro (01-07)Document7 pagesMeta 2.1 Ocampo Libro (01-07)germanNo ratings yet
- Meta 1.1 Ocampo Libro (11-21)Document11 pagesMeta 1.1 Ocampo Libro (11-21)germanNo ratings yet
- Meta 1.1 Ocampo Libro (11-21) (01-06)Document6 pagesMeta 1.1 Ocampo Libro (11-21) (01-06)germanNo ratings yet
- Meta 2.2 Ocampo LibroDocument7 pagesMeta 2.2 Ocampo LibrogermanNo ratings yet
- Carcinogens: Environmental and Genetic FactorsDocument3 pagesCarcinogens: Environmental and Genetic FactorsSyed Ali HassanNo ratings yet
- Mook 2004Document21 pagesMook 2004BerryEkaPardaBancinNo ratings yet
- HistocologyDocument10 pagesHistocologyKelvin HillsNo ratings yet
- LMR 2020Document106 pagesLMR 2020VideoScoNo ratings yet
- Delitto PerfettoDocument6 pagesDelitto Perfettoanirbanmanna88320No ratings yet
- Trusted Medical Answers-In Seconds.: GonorrheaDocument7 pagesTrusted Medical Answers-In Seconds.: GonorrheantnquynhproNo ratings yet
- Total Gut Restoration Patient HandoutDocument2 pagesTotal Gut Restoration Patient HandoutChristianNo ratings yet
- Pre Transfusion TestingDocument67 pagesPre Transfusion TestingPaulino GarciaNo ratings yet
- Tumor Markers: Dr. Hendy Wijaya, M. BiomedDocument23 pagesTumor Markers: Dr. Hendy Wijaya, M. BiomedSeptiansyah CahyoNo ratings yet
- Fcps-1 August 2022 Bcqs by DR Asif Ali Khan (Author of Dod Treatment Guide)Document150 pagesFcps-1 August 2022 Bcqs by DR Asif Ali Khan (Author of Dod Treatment Guide)qwertcccNo ratings yet
- 2012 Number 1 947445146213461Document5 pages2012 Number 1 947445146213461MedstudNo ratings yet
- Mr. Rajiv Ranjan Prasad 30 Yrs/Male 817124 17/09/2020 10:06AM 17/09/2020 11:07AMDocument2 pagesMr. Rajiv Ranjan Prasad 30 Yrs/Male 817124 17/09/2020 10:06AM 17/09/2020 11:07AMRajiv RanjanNo ratings yet
- Dengue by KhushalDocument33 pagesDengue by Khushalikram ullah khanNo ratings yet
- Tonsillitis and Peritonsillar AbscessDocument19 pagesTonsillitis and Peritonsillar AbscesszayNo ratings yet
- Klenow Fragment - Wikipedia, The Free EncyclopediaDocument2 pagesKlenow Fragment - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet
- Peran Vitamin B6 Terhadap Inflamasi Pada Adhesi Peritoneal Pasca Laparotomi: Tinjauan PustakaDocument7 pagesPeran Vitamin B6 Terhadap Inflamasi Pada Adhesi Peritoneal Pasca Laparotomi: Tinjauan PustakaDiBi DBNo ratings yet
- in Response YanDocument13 pagesin Response Yaninappropriate entertainmentNo ratings yet
- Anti-IL-21 Monoclonal Antibody Combined With Liraglutide Effectively Reverses Established Hyperglycemia in Mouse Models of Type 1 Diabetes.Document10 pagesAnti-IL-21 Monoclonal Antibody Combined With Liraglutide Effectively Reverses Established Hyperglycemia in Mouse Models of Type 1 Diabetes.rafaelpires85No ratings yet
- Vibrio Cholerae - Vibrio Mimicus: Bacte Lec Quiz 2Document3 pagesVibrio Cholerae - Vibrio Mimicus: Bacte Lec Quiz 2KENT BENEDICT PERALESNo ratings yet
- 1 2lec General Amoebiasis 3Document11 pages1 2lec General Amoebiasis 3بلسم محمود شاكرNo ratings yet
- (Updated) CHAPTER 18 Haemophilus (HACEK + - Ella)Document49 pages(Updated) CHAPTER 18 Haemophilus (HACEK + - Ella)Nel TinduganiNo ratings yet
- LaryngitisDocument53 pagesLaryngitisEkky Rizky MaulanaNo ratings yet
- Manajemen Hiv.1Document92 pagesManajemen Hiv.1Ariestha Teza AdipratamaNo ratings yet