You might also like
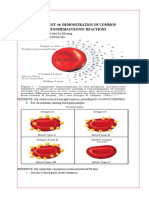
- Experiment #4: Demonstration of Common Immunohematologic ReactionsDocument5 pagesExperiment #4: Demonstration of Common Immunohematologic ReactionsKriziaNo ratings yet
- The Spirit Controlled Life Ebook PDF 1Document184 pagesThe Spirit Controlled Life Ebook PDF 1Okesola AbayomiNo ratings yet
- Synthesis Essay Coming To Grips With GenesisDocument11 pagesSynthesis Essay Coming To Grips With Genesisapi-259381516No ratings yet
- Flow Zone Indicator Guided Workflows For PetrelDocument11 pagesFlow Zone Indicator Guided Workflows For PetrelAiwarikiaar100% (1)
- Carinthia Katalog DownloadDocument16 pagesCarinthia Katalog DownloadOperator_010100% (2)
- Notes Transfer of Thermal EnergyDocument12 pagesNotes Transfer of Thermal Energymahrosh mamoon100% (2)
- Rachel Joyce - A Snow Garden and Other Stories PDFDocument118 pagesRachel Joyce - A Snow Garden and Other Stories PDFИгорь ЯковлевNo ratings yet
- Medical GeneticsDocument4 pagesMedical GeneticsCpopNo ratings yet
- Colour FastnessDocument37 pagesColour FastnessSivakumar K100% (1)
- A Multiscale Red Blood Cell Model With Accurate Mechanics, Rheology, and DynamicsDocument11 pagesA Multiscale Red Blood Cell Model With Accurate Mechanics, Rheology, and DynamicsangelaNo ratings yet
- Eritrocito CtiesqueletoDocument6 pagesEritrocito CtiesqueletodetrNo ratings yet
- 2007 PNAS Cytoskeletal-DynamicsDocument6 pages2007 PNAS Cytoskeletal-DynamicsAdi PasahNo ratings yet
- Journal Pone 0259267Document19 pagesJournal Pone 0259267Coelho FrancoNo ratings yet
- BB BasicsDocument5 pagesBB BasicsRachelle VillanuevaNo ratings yet
- Full Dynamics of A Red Blood Cell in Shear Ow: 20808 - 20813 Pnas December 18, 2012 Vol. 109 No. 51Document6 pagesFull Dynamics of A Red Blood Cell in Shear Ow: 20808 - 20813 Pnas December 18, 2012 Vol. 109 No. 51rezqNo ratings yet
- Modeling of Biomechanics and Biorheology of Red Blood Cells in Type 2 Diabetes MellitusDocument10 pagesModeling of Biomechanics and Biorheology of Red Blood Cells in Type 2 Diabetes Mellitusyour mama123No ratings yet
- Motion of A Tumour Cell Under The Blood Flow at Low Reynolds Number in A Curved MicrovesselDocument10 pagesMotion of A Tumour Cell Under The Blood Flow at Low Reynolds Number in A Curved MicrovesselJuana Valentina CamachoNo ratings yet
- Red Blood Cell MembraneDocument28 pagesRed Blood Cell MembraneAisyah Atikah100% (1)
- Good Blood MicroDocument10 pagesGood Blood MicroAllanNo ratings yet
- An Immersed Boundary-Lattice Boltzmann Model For Simulation of Malaria-Infected Red Blood Cell in Micro-ChannelDocument8 pagesAn Immersed Boundary-Lattice Boltzmann Model For Simulation of Malaria-Infected Red Blood Cell in Micro-ChannelYangNo ratings yet
- Accepted Manuscript: Microvascular ResearchDocument27 pagesAccepted Manuscript: Microvascular ResearchDzigi StarijiNo ratings yet
- Ves 5Document16 pagesVes 5Khushii NaamdeoNo ratings yet
- Paper FSI3 FINAL REVISED MS2Document29 pagesPaper FSI3 FINAL REVISED MS2Amir Reza MohebbiNo ratings yet
- PHY Fluids WBC KineticsDocument22 pagesPHY Fluids WBC KineticsRamesh KavaliNo ratings yet
- Shape Changes of Erythrocytes During Blood Clot Contraction and The Structure of PolyhedrocytesDocument14 pagesShape Changes of Erythrocytes During Blood Clot Contraction and The Structure of Polyhedrocytesjvp jvpNo ratings yet
- Modeling of Spontaneous Penetration of Viscoelastic FluidsDocument10 pagesModeling of Spontaneous Penetration of Viscoelastic Fluidssu67983629No ratings yet
- Link (2020) - A MATHEMATICAL MODEL OF PLATELET AGGREGATION IN AN EXTRAVASCULAR INJURY UNDER FLOW.Document35 pagesLink (2020) - A MATHEMATICAL MODEL OF PLATELET AGGREGATION IN AN EXTRAVASCULAR INJURY UNDER FLOW.Vint PineNo ratings yet
- Discrete Phase Model of Blood Flow in A Roughness Microchannel Simulating The Formation of PseudointimaDocument14 pagesDiscrete Phase Model of Blood Flow in A Roughness Microchannel Simulating The Formation of Pseudointimablack userNo ratings yet
- J. Comput. Theor. Nanosci. 7 (2010) 683-692, Doi:10.1166/jctn.2010.1412Document23 pagesJ. Comput. Theor. Nanosci. 7 (2010) 683-692, Doi:10.1166/jctn.2010.1412Shaikh Saifullah KhalidNo ratings yet
- Computer Methods and Programs in Biomedicine: Arash Mollahosseini, Srija Argumeedi, Amira Abdelrasoul, Ahmed ShokerDocument16 pagesComputer Methods and Programs in Biomedicine: Arash Mollahosseini, Srija Argumeedi, Amira Abdelrasoul, Ahmed ShokerStalwart sheikhNo ratings yet
- BR J Haematol - 2019 - O Donnell - Advances in Understanding The Molecular Mechanisms That Maintain Normal HaemostasisDocument13 pagesBR J Haematol - 2019 - O Donnell - Advances in Understanding The Molecular Mechanisms That Maintain Normal HaemostasisMita AdrianiNo ratings yet
- 2006 Biochimie 88 1229-1239Document11 pages2006 Biochimie 88 1229-1239mbrylinskiNo ratings yet
- Fujiwara 2009Document6 pagesFujiwara 2009Inass ELHAOUARINo ratings yet
- Summary of Research PaperDocument11 pagesSummary of Research PaperNhhjNo ratings yet
- Journal Pone 0174795Document15 pagesJournal Pone 0174795Med Hatem AlloucheNo ratings yet
- Hemoperfusion: Technical Aspects and State of The Art: Review Open AccessDocument12 pagesHemoperfusion: Technical Aspects and State of The Art: Review Open AccessTisaNo ratings yet
- 1 s2.0 S1011134422000525 MainDocument12 pages1 s2.0 S1011134422000525 MainTatianne MatosNo ratings yet
- Van Oss y DlvoDocument15 pagesVan Oss y DlvoaleiviNo ratings yet
- Applied Physics LettersDocument6 pagesApplied Physics LettersChirodeep BakliNo ratings yet
- Mitotic Chromosome Compaction Via Active Loop Extrusion - Biology Et Al. - 2015Document43 pagesMitotic Chromosome Compaction Via Active Loop Extrusion - Biology Et Al. - 2015innu88No ratings yet
- Armstrong 2004Document12 pagesArmstrong 2004aguslistyNo ratings yet
- Hemoperfusión Estado Del ArteDocument12 pagesHemoperfusión Estado Del ArtePaolitha Triana MonroyNo ratings yet
- STPP Berger 2004Document16 pagesSTPP Berger 2004Shailender JosephNo ratings yet
- Formation of 3D Cholesterol Crystals From 2D Nucleation Sites in Lipid Bilayer Membranes: Implications For AtherosclerosisDocument7 pagesFormation of 3D Cholesterol Crystals From 2D Nucleation Sites in Lipid Bilayer Membranes: Implications For AtherosclerosisLuzyaneJaimesOrtizNo ratings yet
- Massoudi 2012Document15 pagesMassoudi 2012azNo ratings yet
- Blood Linear Viscoelasticity by Small Amplitude Oscillatory Flow - Tomaiuolo Carciati Caserta Guido - Rheol Acta 2016Document11 pagesBlood Linear Viscoelasticity by Small Amplitude Oscillatory Flow - Tomaiuolo Carciati Caserta Guido - Rheol Acta 2016Amalia RamunnoNo ratings yet
- Red Blood Cell Shape and Fluctuations: Cytoskeleton Confinement and ATP ActivityDocument12 pagesRed Blood Cell Shape and Fluctuations: Cytoskeleton Confinement and ATP ActivityDewi Sekar MiasihNo ratings yet
- BERGER 2004 - 2 - Rede de Reticulação Quitosana (Cov e Ion)Document16 pagesBERGER 2004 - 2 - Rede de Reticulação Quitosana (Cov e Ion)Jéssica FonsecaNo ratings yet
- Light Effect On Water Viscosity - Implications Fot ATP BiosynthesisDocument6 pagesLight Effect On Water Viscosity - Implications Fot ATP BiosynthesisAlejandro HerreraNo ratings yet
- Rivera Solorio2006Document10 pagesRivera Solorio2006Atrocitus RedNo ratings yet
- Rayleigh Coated FibersDocument14 pagesRayleigh Coated Fibersmexicanu99No ratings yet
- Matsumoto 2007Document10 pagesMatsumoto 2007Trí DũngNo ratings yet
- Measurement Techniques For Red Blood Cell Deformability: Recent AdvancesDocument28 pagesMeasurement Techniques For Red Blood Cell Deformability: Recent AdvancesangelaNo ratings yet
- 5 BD PDFDocument6 pages5 BD PDFHyakuya YuuichiroNo ratings yet
- 205 Cooper2011AngewChemDocument4 pages205 Cooper2011AngewChemDan AlarNo ratings yet
- Rhodes 201998Document7 pagesRhodes 201998انتصار ميلادNo ratings yet
- JSB 142 108 03Document25 pagesJSB 142 108 03Soma GhoshNo ratings yet
- Absorption Spectroscopy of Single Red Blood Cells in The Presence of Mechanical Deformations Induced by Optical TrapsDocument9 pagesAbsorption Spectroscopy of Single Red Blood Cells in The Presence of Mechanical Deformations Induced by Optical Trapsbsk kumarNo ratings yet
- Studying Phase Separation in Confinement: SciencedirectDocument12 pagesStudying Phase Separation in Confinement: SciencedirectJuliana VilachãNo ratings yet
- Self Organisation and Convection of Confined Magnetotactic BacteriaDocument9 pagesSelf Organisation and Convection of Confined Magnetotactic BacteriaPamela MuñozNo ratings yet
- Journal of Molecular LiquidsDocument11 pagesJournal of Molecular Liquidsanon_475646261No ratings yet
- 2020 MONITORING COHERENCE Hemodynamic Coherence and The Rationale For MONITORING PDFDocument13 pages2020 MONITORING COHERENCE Hemodynamic Coherence and The Rationale For MONITORING PDFLUIS CARLOS MAYANo ratings yet
- Plasma Extraction by Centrifugo-PneumatiDocument6 pagesPlasma Extraction by Centrifugo-PneumatihanieNo ratings yet
- Xu - Porolate Compound Capsules - 2020Document9 pagesXu - Porolate Compound Capsules - 2020Zohre AlinejadNo ratings yet
- Rheology For CliniciansDocument8 pagesRheology For CliniciansRaluca LNo ratings yet
- Pharmaceutics 6-v2Document44 pagesPharmaceutics 6-v2Edward NoriegaNo ratings yet
- Potential For Modulation of The Hydrophobic Effect Inside ChaperoninsDocument9 pagesPotential For Modulation of The Hydrophobic Effect Inside Chaperoninsmasteranza5936No ratings yet
- ANG DELAVEGA FinalPositionPaperDocument6 pagesANG DELAVEGA FinalPositionPaperZane IbalaNo ratings yet
- Bravo MRI II Operation ManualDocument45 pagesBravo MRI II Operation ManualLuis100% (1)
- GSD Puppy Training Essentials PDFDocument2 pagesGSD Puppy Training Essentials PDFseja saulNo ratings yet
- Warehouse Management Solution SheetDocument2 pagesWarehouse Management Solution Sheetpatelnandini109No ratings yet
- ARK - Intel® Core™ I3-370m Processor (3M Cache, 2Document3 pagesARK - Intel® Core™ I3-370m Processor (3M Cache, 2Delzi Guindra AdriNo ratings yet
- Chapter 7 Dopant Diffusion - IDocument32 pagesChapter 7 Dopant Diffusion - I강준호No ratings yet
- AS and A Level: ChemistryDocument11 pagesAS and A Level: ChemistryStingy BieNo ratings yet
- Water Filling MachineDocument15 pagesWater Filling Machinepallab D RozarioNo ratings yet
- Patel 2013Document116 pagesPatel 2013hnphuocNo ratings yet
- LinkageDocument9 pagesLinkageHarshu JunghareNo ratings yet
- RD Sharma Class8 SolutionsDocument2 pagesRD Sharma Class8 Solutionsncertsoluitons100% (2)
- Data SiEMEx School SafetyPreparedness 25 26 NOVDocument81 pagesData SiEMEx School SafetyPreparedness 25 26 NOVSuraj RajuNo ratings yet
- ELS 06 Maret 223Document16 pagesELS 06 Maret 223Tri WinarsoNo ratings yet
- Haldex-Barnes 2-Stage Pump For Log SplittersDocument2 pagesHaldex-Barnes 2-Stage Pump For Log SplittersPer Akkamaan AgessonNo ratings yet
- Segmentation of Blood Vessels Using Rule-Based and Machine-Learning-Based Methods: A ReviewDocument10 pagesSegmentation of Blood Vessels Using Rule-Based and Machine-Learning-Based Methods: A ReviewRainata PutraNo ratings yet
- Automatic Train OperationDocument6 pagesAutomatic Train OperationAnupam KhandelwalNo ratings yet
- 331-10 331 Operators Manual enDocument12 pages331-10 331 Operators Manual enYahir VidalNo ratings yet
- Crma Unit 1 Crma RolesDocument34 pagesCrma Unit 1 Crma Rolesumop3plsdn0% (1)
- Case AnalysisDocument2 pagesCase AnalysisJessa San PedroNo ratings yet
- 23001864Document15 pages23001864vinodsrawat33.asiNo ratings yet
- Presentation AcetanilideDocument22 pagesPresentation AcetanilideNovitasarii JufriNo ratings yet
- End of Summer Course Test August 2021Document16 pagesEnd of Summer Course Test August 2021Thanh Nguyen vanNo ratings yet