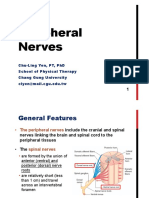
You might also like
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesFrom EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesNo ratings yet
- Nerve Degeneration and Regeneration: A SummaryDocument36 pagesNerve Degeneration and Regeneration: A SummaryPriyanka RameshNo ratings yet
- Pathophysiology of Peripheral Nerve InjuryDocument18 pagesPathophysiology of Peripheral Nerve InjuryshreyasNo ratings yet
- Degeneration and Regeneration of Nerve FibreDocument25 pagesDegeneration and Regeneration of Nerve Fibresunil patelNo ratings yet
- Degeneration & Regeneration of Nerve FibreDocument29 pagesDegeneration & Regeneration of Nerve FibreSana Fatima100% (2)
- Degeneration & Regeneration of Nerve FibreDocument29 pagesDegeneration & Regeneration of Nerve FibreSana FatimaNo ratings yet
- Wallerian DegenerationqDocument8 pagesWallerian DegenerationqNST MtzNo ratings yet
- Regeneration and Repair of Peripheral Nerves: Elizabeth O. Johnson, Aristides B. Zoubos, Panayotis N. SoucacosDocument6 pagesRegeneration and Repair of Peripheral Nerves: Elizabeth O. Johnson, Aristides B. Zoubos, Panayotis N. SoucacosFrank De'doctorNo ratings yet
- Unit 2 NeuroplasticityDocument6 pagesUnit 2 NeuroplasticityKhushi AroraNo ratings yet
- Nerve InjuryDocument17 pagesNerve InjuryachalsinghbdnNo ratings yet
- Mood disorderDocument5 pagesMood disorderAthulya MkNo ratings yet
- NeuroradiologyCurrent and Future Imaging of The Peripheral NervoussystemDocument10 pagesNeuroradiologyCurrent and Future Imaging of The Peripheral NervoussystemRyanNo ratings yet
- 15.09.14. Pro. Nerve Fiber TypesDocument33 pages15.09.14. Pro. Nerve Fiber Typesashok_solanki_23No ratings yet
- Regenerasi Saraf Perifer (Peripheral Nerve Regeneration) : December 2014Document12 pagesRegenerasi Saraf Perifer (Peripheral Nerve Regeneration) : December 2014ferryNo ratings yet
- Nerve RegenDocument31 pagesNerve RegenKAPIL SINGHNo ratings yet
- Study Guide-Regeneration and RecoveryDocument2 pagesStudy Guide-Regeneration and RecoveryjoeyNo ratings yet
- Peripheral Nerve Injury and Repair: Adam Osbourne, 5th Year MedicineDocument5 pagesPeripheral Nerve Injury and Repair: Adam Osbourne, 5th Year Medicineayrahma29No ratings yet
- Pathophysiology of Nerve: Dr. Ayisha Qureshi Professor & Hod MBBS, MphilDocument26 pagesPathophysiology of Nerve: Dr. Ayisha Qureshi Professor & Hod MBBS, MphilMuhammad AhmedNo ratings yet
- Development of Nervous SystemDocument71 pagesDevelopment of Nervous SystemAbdimalik AliNo ratings yet
- ANA 221 - Reticular Formation, Nerve Re DegenerationDocument18 pagesANA 221 - Reticular Formation, Nerve Re DegenerationOtoto ForexNo ratings yet
- Nerve Muscle PhysioDocument30 pagesNerve Muscle Physiopatel_hanisha06No ratings yet
- PT Pediatric Surgery Sec 1Document27 pagesPT Pediatric Surgery Sec 1Amany SaifNo ratings yet
- Neuroplasticity - Chap 6Document3 pagesNeuroplasticity - Chap 6Althea YabutNo ratings yet
- V. Pathophysiology: Nerve InjuryDocument4 pagesV. Pathophysiology: Nerve Injuryjoanna gurtizaNo ratings yet
- Degeneration and Regenration of Nerve Fibers by Dr. RoomiDocument8 pagesDegeneration and Regenration of Nerve Fibers by Dr. RoomiMudassar Roomi100% (1)
- Biomechanics of Nerve 211080006 Rehab AymanDocument9 pagesBiomechanics of Nerve 211080006 Rehab Aymanrehab aymanNo ratings yet
- Anaphy Batt CutieeeDocument3 pagesAnaphy Batt Cutieee매레이디No ratings yet
- Gingival Mesenchymal Stem Cells, Concentrated Growth Factors and Silk-Fibroin Scaffold To Alleviate Peripheral Nerve Regeneration: A ReviewDocument6 pagesGingival Mesenchymal Stem Cells, Concentrated Growth Factors and Silk-Fibroin Scaffold To Alleviate Peripheral Nerve Regeneration: A ReviewGhinalya Chalbi AnandaNo ratings yet
- Nerve Healing FinalDocument31 pagesNerve Healing FinalKim MateoNo ratings yet
- Peripheral Nerve Injury 2Document24 pagesPeripheral Nerve Injury 2Gargi MPNo ratings yet
- Nervous and Reproductive Note.Document45 pagesNervous and Reproductive Note.Sushil AcharyaNo ratings yet
- Nerve-Part 1Document30 pagesNerve-Part 1GiftNo ratings yet
- Epineurium. If The Nerve Contains More Than One Fascicle, The Epineurium May Be Divided IntoDocument18 pagesEpineurium. If The Nerve Contains More Than One Fascicle, The Epineurium May Be Divided Intoayaanshariq99No ratings yet
- ChinJPhysiol664209-1981182 053011Document11 pagesChinJPhysiol664209-1981182 053011Ana Sofia SantiagoNo ratings yet
- Jurnal Reading Ortho 1Document6 pagesJurnal Reading Ortho 1midellialintinNo ratings yet
- Spinal Cord InjuryDocument19 pagesSpinal Cord Injuryprof01faridNo ratings yet
- Henne Man 1965Document21 pagesHenne Man 1965Brayan BuitragoNo ratings yet
- Degeneration and Regenration of Nerve Fibers by Dr. RoomiDocument11 pagesDegeneration and Regenration of Nerve Fibers by Dr. RoomiMudassar RoomiNo ratings yet
- Unit III Development and Plasticity of The BrainDocument10 pagesUnit III Development and Plasticity of The Brainjusq marimarNo ratings yet
- Neuro Exam 3Document17 pagesNeuro Exam 3John RobinsonNo ratings yet
- The History of Nerve RepairDocument32 pagesThe History of Nerve RepairWim ZhuNo ratings yet
- Group 9 Neuron Structure & FunctionDocument14 pagesGroup 9 Neuron Structure & FunctionMosesNo ratings yet
- NeuropathologyDocument66 pagesNeuropathologyKaramsi Gopinath NaikNo ratings yet
- Option A - Neurobiology and BehaviourDocument29 pagesOption A - Neurobiology and Behaviour玛丽娜No ratings yet
- Peripheral Nerve InjuriesDocument12 pagesPeripheral Nerve InjuriesJiggs LimNo ratings yet
- Week 5 Peripheral NervesDocument32 pagesWeek 5 Peripheral Nervesray RayNo ratings yet
- Gordon 2016Document16 pagesGordon 2016Indah GitaswariNo ratings yet
- Spinal Cord InjuryDocument17 pagesSpinal Cord InjuryMuhammad FaridNo ratings yet
- Neural stem cell-derived extracellular vesicles favour neuronal differentiation and plasticity under stress conditionsDocument10 pagesNeural stem cell-derived extracellular vesicles favour neuronal differentiation and plasticity under stress conditionsGedguol AlvarezNo ratings yet
- Nerve Injury Repair and Healing GuideDocument34 pagesNerve Injury Repair and Healing GuideAtikah PurnamasariNo ratings yet
- Macrophage-Induced Blood Vessels Guide Schwann Cell RegenerationDocument14 pagesMacrophage-Induced Blood Vessels Guide Schwann Cell RegenerationDeng PanNo ratings yet
- Development of The Central Nervous SystemDocument39 pagesDevelopment of The Central Nervous Systemkutticute_877110165No ratings yet
- 1.3 Neural Development and Neurogenesis (Final)Document4 pages1.3 Neural Development and Neurogenesis (Final)Austin CabreraNo ratings yet
- Nerve InjuryDocument7 pagesNerve Injuryzwecker4458No ratings yet
- Midtern LabDocument21 pagesMidtern LabJustine Mae OyongNo ratings yet
- Module I DevPsychDocument5 pagesModule I DevPsychsammparker090297No ratings yet
- Binary Fission Process & Examples in Protists & TapewormsDocument24 pagesBinary Fission Process & Examples in Protists & Tapewormsbanapte599706No ratings yet
- Neuro anatomy, motor neuronDocument9 pagesNeuro anatomy, motor neuronMichele Silva PiresNo ratings yet
- Nerve Injury Classification, Nerve Degeneration, Regeneration and Repair TechniquesDocument50 pagesNerve Injury Classification, Nerve Degeneration, Regeneration and Repair TechniquesRajveerNo ratings yet
- Myelination and Regional Domain Differentiation of The Axon.Document25 pagesMyelination and Regional Domain Differentiation of The Axon.Thais MtNo ratings yet
- BridgewaterPureAlpha CaseStudy MPIDocument9 pagesBridgewaterPureAlpha CaseStudy MPIMichael Guan100% (2)
- Ong v. BognalbalDocument2 pagesOng v. BognalbalStefanRodriguez100% (3)
- Pfe Training AvDocument20 pagesPfe Training AvpankajNo ratings yet
- A Literature Survey On The Accuracy of Software Effort Estimation ModelsDocument6 pagesA Literature Survey On The Accuracy of Software Effort Estimation Modelsutkarsh sharmaNo ratings yet
- Term Paper of Operating SystemDocument25 pagesTerm Paper of Operating Systemnannupriya100% (1)
- Prescription AssignmentDocument6 pagesPrescription AssignmentChiela Alcantara BagnesNo ratings yet
- Finding the Common Point of Convex SetsDocument18 pagesFinding the Common Point of Convex SetsAditya PillaiNo ratings yet
- Hughie's Financial StatusDocument3 pagesHughie's Financial StatusGuna SeelanNo ratings yet
- Australian Guidebook For Structural Engineers by Lonnie Pack 1138031852Document5 pagesAustralian Guidebook For Structural Engineers by Lonnie Pack 1138031852Frans ChandraNo ratings yet
- Diff BTW CV and ResumeDocument3 pagesDiff BTW CV and ResumeHabib Ullah Habib UllahNo ratings yet
- Argumentative Essay - Sample 2 - Students Should Spend Less Time Listening To MusicDocument2 pagesArgumentative Essay - Sample 2 - Students Should Spend Less Time Listening To MusicRidhwan AfiffNo ratings yet
- The Authors Guild vs. Google, Inc. Facts: Plaintiffs, Who Are Authors of Published Books UnderDocument15 pagesThe Authors Guild vs. Google, Inc. Facts: Plaintiffs, Who Are Authors of Published Books UnderElead Gaddiel S. AlbueroNo ratings yet
- Exalted Glories of The Most High - The Maidens of Destiny 2eDocument39 pagesExalted Glories of The Most High - The Maidens of Destiny 2egercog950% (1)
- Proactive Network Defence 7CS017Document15 pagesProactive Network Defence 7CS017Anchal SinghNo ratings yet
- Radar Engineering, 1/e: Book Information Sheet Book Information SheetDocument2 pagesRadar Engineering, 1/e: Book Information Sheet Book Information SheetLakshmi MounikaNo ratings yet
- Research in Daily Life 2Document166 pagesResearch in Daily Life 2Zianna Marie De RamosNo ratings yet
- Journal All Pages Jan2019 PDFDocument68 pagesJournal All Pages Jan2019 PDFmahadev_prasad_7No ratings yet
- Reflection On Teaching Practice To YLsDocument3 pagesReflection On Teaching Practice To YLsivannasoledadNo ratings yet
- Extraction of Rare Earth Oxides From Discarded Compact - 2019 - Minerals EngineDocument10 pagesExtraction of Rare Earth Oxides From Discarded Compact - 2019 - Minerals EnginepHNo ratings yet
- Astrology Certificate Syllabus EnglishDocument4 pagesAstrology Certificate Syllabus EnglishDominic SavioNo ratings yet
- Professional CV FormatDocument2 pagesProfessional CV FormatShawn ParkerNo ratings yet
- Jihvamuliya UpadhmaniyaDocument6 pagesJihvamuliya UpadhmaniyaPradeepa SerasingheNo ratings yet
- DSC MatDocument41 pagesDSC MatRamuCivilNo ratings yet
- Mental Health and Self-Care ReflectionDocument2 pagesMental Health and Self-Care Reflectionapi-472652222No ratings yet
- NIH Public Access: Sleep and Women's HealthDocument31 pagesNIH Public Access: Sleep and Women's HealthHusni FaridNo ratings yet
- OBATALA - Ifa and The Chief of The Spirit of The White ClothDocument5 pagesOBATALA - Ifa and The Chief of The Spirit of The White ClothToyin Adepoju100% (3)
- History of Indian CinemaDocument14 pagesHistory of Indian Cinemaswatikuk100% (9)
- Radar Clutter Types and Models ExplainedDocument63 pagesRadar Clutter Types and Models ExplainedWesley GeorgeNo ratings yet
- Chapter 2-Meaning and Scope of Public FinanceDocument21 pagesChapter 2-Meaning and Scope of Public FinanceyebegashetNo ratings yet
- Feminine Beauty, National Identity and Political Conflict in Postwar ItalyDocument21 pagesFeminine Beauty, National Identity and Political Conflict in Postwar ItalyHugyecz DorinaNo ratings yet