You might also like
- Celery Juice Miracle JuiceDocument16 pagesCelery Juice Miracle JuiceAntonio Prata96% (54)
- Integrated Cytology of Cerebrospinal FluidDocument93 pagesIntegrated Cytology of Cerebrospinal FluidVishal Maindarkar100% (1)
- The-Patient-Perspective - What-POTS Patients Want Physicians To KnowDocument5 pagesThe-Patient-Perspective - What-POTS Patients Want Physicians To KnowEmilie ChateletNo ratings yet
- Core Curriculum For Transplant Nurses 2nd Edition Ebook PDFDocument61 pagesCore Curriculum For Transplant Nurses 2nd Edition Ebook PDFmichael.quinonez375100% (33)
- POTS and Other Forms or Orthostatic Intolerance in Ehlers-Danlos SyndromeDocument8 pagesPOTS and Other Forms or Orthostatic Intolerance in Ehlers-Danlos SyndromeEmilie ChateletNo ratings yet
- Chapter 8 Infectious Diseases Robbins and Cotran Pathologic Basis of Disease PDFDocument12 pagesChapter 8 Infectious Diseases Robbins and Cotran Pathologic Basis of Disease PDFRitz Celso100% (1)
- MUST To KNOW in Mycology and VirologyDocument18 pagesMUST To KNOW in Mycology and VirologyCaraflos Pangilinan67% (3)
- The Bread Baker's Apprentice Bagels - V-SpotDocument9 pagesThe Bread Baker's Apprentice Bagels - V-SpotEmilie ChateletNo ratings yet
- The Role of Edible Mushrooms in Health Evaluation of The EvidenceDocument23 pagesThe Role of Edible Mushrooms in Health Evaluation of The EvidencekierkegardNo ratings yet
- 先天免疫学糖生物学Document673 pages先天免疫学糖生物学Wenbo Yan100% (1)
- Infectious MononucleosisDocument35 pagesInfectious MononucleosisShv Naga100% (1)
- Summary of Recent Significant Findings in ME-CFS Research - Updated February 2019 PDFDocument21 pagesSummary of Recent Significant Findings in ME-CFS Research - Updated February 2019 PDFEmilie ChateletNo ratings yet
- Chapter 24 DNA Viruses That Infect HumansDocument64 pagesChapter 24 DNA Viruses That Infect HumansJJ AlmagroNo ratings yet
- Tartine Bread Bakery Reveals Its SecretsDocument3 pagesTartine Bread Bakery Reveals Its SecretsEmilie ChateletNo ratings yet
- MICROBILOGYDocument13 pagesMICROBILOGYDjdjjd SiisusNo ratings yet
- Carlberg C. Molecular Immunology. How Science Works 2022Document228 pagesCarlberg C. Molecular Immunology. How Science Works 2022Philip Rodríguez GarcíaNo ratings yet
- ImmunologyDocument83 pagesImmunologyKühne ZoeNo ratings yet
- Fundamental Techniques in Cell CultureDocument64 pagesFundamental Techniques in Cell Cultureifti007No ratings yet
- 2015 Immunology Whitebook PDFDocument89 pages2015 Immunology Whitebook PDFZhraa FarisNo ratings yet
- IchthyosisDocument154 pagesIchthyosisprajnamitaNo ratings yet
- Guillain-Barr?? Syndrome: Epidemiology, Pathophysiology and ManagementDocument15 pagesGuillain-Barr?? Syndrome: Epidemiology, Pathophysiology and Managementshf.mxlikNo ratings yet
- Guillenbare 3Document15 pagesGuillenbare 3indadzilarsyNo ratings yet
- Ingles 4 Tesis Abayomi SorinolaDocument154 pagesIngles 4 Tesis Abayomi SorinolaThe Almighty BelugaNo ratings yet
- Guillain-Barr e Syndrome: Epidemiology, Pathophysiology and ManagementDocument15 pagesGuillain-Barr e Syndrome: Epidemiology, Pathophysiology and ManagementErica BettsNo ratings yet
- Inmuno S7Document6 pagesInmuno S7Daniela Fdz CNo ratings yet
- 2016 - Revisão EMDocument9 pages2016 - Revisão EMilc67123No ratings yet
- Fisher Syndrome and Bickerstaff Brainstem Encephalitis (Fisher-Bickerstaff Syndrome)Document9 pagesFisher Syndrome and Bickerstaff Brainstem Encephalitis (Fisher-Bickerstaff Syndrome)John AnastasiadesNo ratings yet
- Cannabinoides Induce Apoptosis en Celulas TDocument8 pagesCannabinoides Induce Apoptosis en Celulas TjgcmNo ratings yet
- Aneuploidy and TumorigenesisDocument7 pagesAneuploidy and TumorigenesisAlejandraOsegueraNo ratings yet
- Key Concepts in Immunology: VaccineDocument12 pagesKey Concepts in Immunology: VaccineChun Hao TheoNo ratings yet
- Acute Bacterial Meningitis in Infants and Children: Paediatric Drugs December 2011Document17 pagesAcute Bacterial Meningitis in Infants and Children: Paediatric Drugs December 2011indahrizkygNo ratings yet
- Microbial Pathogenesis: Nidhi Chauhan, Sukirti Tiwari, Utkarsh JainDocument9 pagesMicrobial Pathogenesis: Nidhi Chauhan, Sukirti Tiwari, Utkarsh JainShofi Dhia AiniNo ratings yet
- Modulación de Los AdrenocorticoidesDocument55 pagesModulación de Los AdrenocorticoidesGabrielaNo ratings yet
- Ebiomedicine: Karis Little, Jacey H. Ma, Nan Yang, Mei Chen, Heping XuDocument9 pagesEbiomedicine: Karis Little, Jacey H. Ma, Nan Yang, Mei Chen, Heping XuSantos Pardo GómezNo ratings yet
- Stem Cell Therapy 1Document6 pagesStem Cell Therapy 1Amit MaityNo ratings yet
- Clinical Microbiology Reviews-2014-Ahmed-631.fullDocument16 pagesClinical Microbiology Reviews-2014-Ahmed-631.fullduverney.gaviriaNo ratings yet
- 72nd 2011 MercuryDocument81 pages72nd 2011 MercurySusantika MurtiniNo ratings yet
- 2009 - Goldberg - Infection and Type 1 Diabetes Mellitus - A Two Edged SwordDocument5 pages2009 - Goldberg - Infection and Type 1 Diabetes Mellitus - A Two Edged SwordHovhannes SahakyanNo ratings yet
- The Bovine Spleen - Interactions Among Splenic Cell Populations in The Innate Immunologic Control of Hemoparasitic InfectionsDocument14 pagesThe Bovine Spleen - Interactions Among Splenic Cell Populations in The Innate Immunologic Control of Hemoparasitic Infectionsmafiajalanan.grup7No ratings yet
- Drug Development Against Tuberculosis Impact of AlkaloidsDocument41 pagesDrug Development Against Tuberculosis Impact of Alkaloidsamine ben dahsenNo ratings yet
- Herpes Simplex Keratitis: Stephen Kaye, Anshoo ChoudharyDocument26 pagesHerpes Simplex Keratitis: Stephen Kaye, Anshoo ChoudharyRafiqy Sa'adiy FaizunNo ratings yet
- Chiang 2016Document313 pagesChiang 2016Anil KumarNo ratings yet
- Physics Reports: Grégoire Altan-Bonnet, Thierry Mora, Aleksandra M. WalczakDocument83 pagesPhysics Reports: Grégoire Altan-Bonnet, Thierry Mora, Aleksandra M. Walczakadi63No ratings yet
- Julyanto Putra Admaja (p1337420921185) Kelas Palangka RayaDocument43 pagesJulyanto Putra Admaja (p1337420921185) Kelas Palangka RayaJULYANTO PUTRA ADMAJANo ratings yet
- Brain, Behavior, and Immunity: Aravinthan Varatharaj, Ian GaleaDocument12 pagesBrain, Behavior, and Immunity: Aravinthan Varatharaj, Ian GaleaDavid James PNo ratings yet
- 1 s2.0 S0956566312006513 MainDocument11 pages1 s2.0 S0956566312006513 Mainbrisj34No ratings yet
- Jurnal Stapilokokus OsteomielitisDocument27 pagesJurnal Stapilokokus OsteomielitisWahyuNo ratings yet
- 1 s2.0 S1568997213000360 MainDocument12 pages1 s2.0 S1568997213000360 MainValiant CheungNo ratings yet
- Blood CompatibilityDocument9 pagesBlood CompatibilitySUYASH SHARMANo ratings yet
- Progress in Neuro-Psychopharmacology & Biological PsychiatryDocument10 pagesProgress in Neuro-Psychopharmacology & Biological Psychiatrynermal93No ratings yet
- Journal of Oral and Maxillofacial Surgery, Medicine, and PathologyDocument9 pagesJournal of Oral and Maxillofacial Surgery, Medicine, and PathologyabdulrazaqNo ratings yet
- Art 3Document26 pagesArt 3renirahmatNo ratings yet
- Antiviral Research 96 (2012) 391-404Document14 pagesAntiviral Research 96 (2012) 391-404mayra alejandra pinilla guerreroNo ratings yet
- Acinetobacter RevisaoDocument45 pagesAcinetobacter RevisaoDaniel Wagner SantosNo ratings yet
- Buccal Micronucleus Cytome AssayDocument14 pagesBuccal Micronucleus Cytome AssayMelannye Caro ManzanoNo ratings yet
- LN Med Bact Final PDFDocument444 pagesLN Med Bact Final PDFSurajNo ratings yet
- LN Med Bact Final PDFDocument444 pagesLN Med Bact Final PDFPrince HimansuNo ratings yet
- 1994 Purpura Trombocitopenica AutoinmuneDocument13 pages1994 Purpura Trombocitopenica AutoinmuneConrado Vera PinedaNo ratings yet
- Meropenem: A Review of Its Use in Patients in Intensive CareDocument28 pagesMeropenem: A Review of Its Use in Patients in Intensive CareHAIDAR RACHMANNo ratings yet
- Progress in Neurobiology: Roger A. Barker, Inez de BeaufortDocument11 pagesProgress in Neurobiology: Roger A. Barker, Inez de BeaufortReemNo ratings yet
- The Immunoregulatory Function of Polyphenols - Implications in Cancer ImmunityDocument11 pagesThe Immunoregulatory Function of Polyphenols - Implications in Cancer ImmunitySusan OstosNo ratings yet
- Evolution of Marek's Disease Virus Pathogenesis and Vaccine ResistanceDocument134 pagesEvolution of Marek's Disease Virus Pathogenesis and Vaccine ResistancedrfaresNo ratings yet
- The Genetics of Human Autoimmune Disease - A Perspective On Progress in The Field and Future Directions Michael F. SeldinDocument12 pagesThe Genetics of Human Autoimmune Disease - A Perspective On Progress in The Field and Future Directions Michael F. Seldinsalwa atsilahNo ratings yet
- Microbiological ResearchDocument7 pagesMicrobiological ResearchDEA REGGIANANo ratings yet
- Antiviral Research: Julie Blaising, Stephen J. Polyak, Eve-Isabelle PécheurDocument11 pagesAntiviral Research: Julie Blaising, Stephen J. Polyak, Eve-Isabelle Pécheurspiridon_andrei2011No ratings yet
- Pti y mm2Document7 pagesPti y mm2Clara Espinosa ArnandisNo ratings yet
- Cockayne Syndrome Clinical Features Source Elsevier SO 2017 (GGT904768966)Document15 pagesCockayne Syndrome Clinical Features Source Elsevier SO 2017 (GGT904768966)dai shujuanNo ratings yet
- (Christophe Boete) Genetically Modified MosquitoesDocument190 pages(Christophe Boete) Genetically Modified Mosquitoesclaudia lilianaNo ratings yet
- Arn MessagerDocument14 pagesArn MessagerMackenzy DrewNo ratings yet
- Endothelial Cells From Innocent Bystanders To Active Par - 2017 - AutoimmunityDocument12 pagesEndothelial Cells From Innocent Bystanders To Active Par - 2017 - AutoimmunityBelia Bima NafisaNo ratings yet
- Adaptive Cellular Stress Pathways As Therapeutic Targets of Dietary Phytochemicals Focus On The Nervous System 2014Document54 pagesAdaptive Cellular Stress Pathways As Therapeutic Targets of Dietary Phytochemicals Focus On The Nervous System 2014upadhyaysk97No ratings yet
- (Prof. Dr. Klaus Wolf (Auth.) ) Nonconventional Yea (B-Ok - Xyz)Document630 pages(Prof. Dr. Klaus Wolf (Auth.) ) Nonconventional Yea (B-Ok - Xyz)Muhammad HusseinNo ratings yet
- Chronic Fatigue Syndrome, Viruses, and The Innate Immune System - Psychiatry AdvisorDocument13 pagesChronic Fatigue Syndrome, Viruses, and The Innate Immune System - Psychiatry AdvisorEmilie ChateletNo ratings yet
- 2022 Conference - Register Now! - International Association For Chronic Fatigue Syndrome - Myalgic Encephalomyelitis (IACFS - ME) PDFDocument6 pages2022 Conference - Register Now! - International Association For Chronic Fatigue Syndrome - Myalgic Encephalomyelitis (IACFS - ME) PDFEmilie ChateletNo ratings yet
- Findings From Critical Illness To Explain Myalgic EncephalomyelitisChronic Fatigue SyndromeDocument13 pagesFindings From Critical Illness To Explain Myalgic EncephalomyelitisChronic Fatigue SyndromeEmilie ChateletNo ratings yet
- MECFS Clinician Coalition Treatment Recs V1Document9 pagesMECFS Clinician Coalition Treatment Recs V1Emilie ChateletNo ratings yet
- HANSON Plasma Metabolomics Reveals Disrupted ResponseDocument56 pagesHANSON Plasma Metabolomics Reveals Disrupted ResponseEmilie ChateletNo ratings yet
- POTS Diagnosis, Physiology 2018 Autonomic NeurosciencesDocument9 pagesPOTS Diagnosis, Physiology 2018 Autonomic NeurosciencesEmilie ChateletNo ratings yet
- The-Gastrointestinal-Symptoms-Present-In-Patients-With POTSDocument8 pagesThe-Gastrointestinal-Symptoms-Present-In-Patients-With POTSEmilie ChateletNo ratings yet
- RELEVANT SCIENTIFIC PUBLICATIONS ON MECFS From 2015-2019Document41 pagesRELEVANT SCIENTIFIC PUBLICATIONS ON MECFS From 2015-2019Emilie ChateletNo ratings yet
- Autonomic Neuroscience: Basic and Clinical: SciencedirectDocument2 pagesAutonomic Neuroscience: Basic and Clinical: SciencedirectEmilie ChateletNo ratings yet
- Brother 2340DW Printer ManualDocument1 pageBrother 2340DW Printer ManualEmilie ChateletNo ratings yet
- How To Enjoy The Zojirushi Home Bakery JuniorDocument30 pagesHow To Enjoy The Zojirushi Home Bakery JuniormilesdzynNo ratings yet
- Chefs Choic WAFFLE PRO 840B MANUALDocument16 pagesChefs Choic WAFFLE PRO 840B MANUALEmilie ChateletNo ratings yet
- 7 Ways To Invite Gratitude, Comfort & Warmth Into Your HomeDocument5 pages7 Ways To Invite Gratitude, Comfort & Warmth Into Your HomeEmilie ChateletNo ratings yet
- WHOLE WHEAT BREAD AND VARATIONS Peter ReinhartDocument9 pagesWHOLE WHEAT BREAD AND VARATIONS Peter ReinhartEmilie ChateletNo ratings yet
- 6 Ways To Love Your Body On Bad Days With A Chronic IllnessDocument12 pages6 Ways To Love Your Body On Bad Days With A Chronic IllnessEmilie ChateletNo ratings yet
- Four Things To Do Every Day For Your Mental HealthDocument5 pagesFour Things To Do Every Day For Your Mental HealthEmilie ChateletNo ratings yet
- Beth Hensperger - The Bread Lover's Bread MachineDocument362 pagesBeth Hensperger - The Bread Lover's Bread MachineEmilie ChateletNo ratings yet
- Reinhart Bread Is Transformational Food On Four Different LevelsDocument1 pageReinhart Bread Is Transformational Food On Four Different LevelsEmilie ChateletNo ratings yet
- 016 13 Hodgkin LymphomaDocument22 pages016 13 Hodgkin LymphomaQuique Zamora MarinNo ratings yet
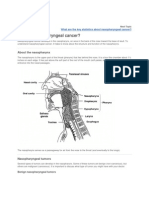
- Nasopharyngeal CancerDocument2 pagesNasopharyngeal CancerCharo FloresNo ratings yet
- Epstein-Barr Virus Super FinalDocument6 pagesEpstein-Barr Virus Super FinalStill DollNo ratings yet
- Autism - Kopie PDFDocument214 pagesAutism - Kopie PDFMohrscribdNo ratings yet
- Epstein-Barr Virus - Associated Lymphomas: Ewelina Grywalska and Jacek RolinskiDocument13 pagesEpstein-Barr Virus - Associated Lymphomas: Ewelina Grywalska and Jacek RolinskiR. FortalNo ratings yet
- Clinical Virology (Dental)Document134 pagesClinical Virology (Dental)samar yousif mohamedNo ratings yet
- Sauropus Androgynus Leaves For Health Benefits: Hype and The ScienceDocument9 pagesSauropus Androgynus Leaves For Health Benefits: Hype and The ScienceRusdiah DiahhNo ratings yet
- Antituberculosis Drug Associated DRESS A Case SeriesDocument8 pagesAntituberculosis Drug Associated DRESS A Case SeriesVivi MaifitrizaNo ratings yet
- Catalogue AMS A4 BD PapDocument29 pagesCatalogue AMS A4 BD PapHienNo ratings yet
- The Spectrum of Antecedent Infections inDocument7 pagesThe Spectrum of Antecedent Infections inmdkev llaalNo ratings yet
- An Unusual Case of Burkitt's Lymphoma Presenting As A Gingival EnlargementDocument7 pagesAn Unusual Case of Burkitt's Lymphoma Presenting As A Gingival Enlargementpoi09No ratings yet
- Herpesviruses, Poxviruses & Human Papilloma Virus: Fort SalvadorDocument33 pagesHerpesviruses, Poxviruses & Human Papilloma Virus: Fort SalvadorFort SalvadorNo ratings yet
- HERPESVIRUSES, PARVO 2021 Students - KopieDocument38 pagesHERPESVIRUSES, PARVO 2021 Students - KopieMr.FantasthiccNo ratings yet
- Human Herpes VirusesDocument40 pagesHuman Herpes Virusesfiea241089No ratings yet
- Transfer Factor-Myths and FactsDocument10 pagesTransfer Factor-Myths and FactsIris BetancourtNo ratings yet
- Hodgkin Lympho PDFDocument371 pagesHodgkin Lympho PDFBertramVrellingNo ratings yet
- Neoplasia: NomenclatureDocument26 pagesNeoplasia: NomenclatureAbdul Samad NoonariNo ratings yet
- Chorus Kit List Rev.03 enDocument6 pagesChorus Kit List Rev.03 enHaitham Salama GhareebNo ratings yet
- Carcinogenic AgentsDocument46 pagesCarcinogenic AgentsSalmanu Labaran ZuruNo ratings yet
- Neoplasia 6: Dr. Eman Krieshan, M.D. 5-1-2022Document34 pagesNeoplasia 6: Dr. Eman Krieshan, M.D. 5-1-2022MOHA BANATNo ratings yet
- Jurnal Karsinoma Nasofaring Dan Infeksi EBV 239-454-1-SMDocument10 pagesJurnal Karsinoma Nasofaring Dan Infeksi EBV 239-454-1-SMHanifahNo ratings yet
- Diseases of The CallitrichidaeDocument48 pagesDiseases of The CallitrichidaeCristhian HernandezNo ratings yet