You might also like
- The Role of 5-HT Systems on Memory and Dysfunctional Memory: Emergent Targets for Memory Formation and Memory AlterationsFrom EverandThe Role of 5-HT Systems on Memory and Dysfunctional Memory: Emergent Targets for Memory Formation and Memory AlterationsNo ratings yet
- The Expanded Biology of SerotoninDocument15 pagesThe Expanded Biology of SerotoninLuiza FonteNo ratings yet
- FCVM 04 00048Document11 pagesFCVM 04 00048Sebastian WallotNo ratings yet
- Serotonin and Molecular Neuroimaging in Humans Using PET PDFDocument19 pagesSerotonin and Molecular Neuroimaging in Humans Using PET PDFWynn TheinNo ratings yet
- Serotonin: Dorotea Mück - (Eler Nela PivacDocument13 pagesSerotonin: Dorotea Mück - (Eler Nela PivacEldwin ChandraNo ratings yet
- Serotonin Systems PDFDocument66 pagesSerotonin Systems PDFdsekulic_1No ratings yet
- Serotonin: A Review: 2008 The Authors. Journal Compilation 2008 Blackwell Publishing LTD 187Document13 pagesSerotonin: A Review: 2008 The Authors. Journal Compilation 2008 Blackwell Publishing LTD 187Lucia LopezNo ratings yet
- Taiwo SeminarDocument48 pagesTaiwo SeminarBishop OlugbengaNo ratings yet
- Hormonii Tiroidieni Și SerotoninaDocument17 pagesHormonii Tiroidieni Și SerotoninaxsmileNo ratings yet
- PIIS0006322398001395 The Spectrum of Behaviors Influenced by SerotoninDocument12 pagesPIIS0006322398001395 The Spectrum of Behaviors Influenced by SerotoninRifqi Hamdani PasaribuNo ratings yet
- BDNF & Skizofrenia Neurodevelopment Nieto 2013Document11 pagesBDNF & Skizofrenia Neurodevelopment Nieto 2013haykal fathirrahmanNo ratings yet
- 5-HTT Independent Effects of Fluoxetine On NeuroplasticityDocument11 pages5-HTT Independent Effects of Fluoxetine On NeuroplasticityCony GSNo ratings yet
- Autism Spectrum Disorder and The Role of SerotoninDocument2 pagesAutism Spectrum Disorder and The Role of SerotoninInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- En 2014-1079Document12 pagesEn 2014-1079Kerin ArdyNo ratings yet
- Serotonin 122Document36 pagesSerotonin 122lepro145100% (2)
- Unit 2 B Viewpoints For Understanding The Causes of Abnormal Behaviour Biological ViewpointDocument22 pagesUnit 2 B Viewpoints For Understanding The Causes of Abnormal Behaviour Biological ViewpointRadhey SurveNo ratings yet
- Accepted Manuscript: Neurobiology of Learning and MemoryDocument49 pagesAccepted Manuscript: Neurobiology of Learning and MemoryChristine HandayaniNo ratings yet
- Hariri2006 PDFDocument10 pagesHariri2006 PDFAhmet Fatih SarıkayaNo ratings yet
- Serotonin & MigraineDocument10 pagesSerotonin & Migrainepetri_jvNo ratings yet
- Neurotransmitters and Psychiatry PDFDocument22 pagesNeurotransmitters and Psychiatry PDFcapriciousbelal93% (14)
- Li Et Al 2017Document8 pagesLi Et Al 2017Ezequiel NascimentoNo ratings yet
- 2011 - Hutchenson, Et Al - Serotonin Receptors and Valve Heart Disease - It Was Meant 2BDocument29 pages2011 - Hutchenson, Et Al - Serotonin Receptors and Valve Heart Disease - It Was Meant 2Bcolompar80No ratings yet
- Li-2020-GABAergic Neurons in The Dorsal Raphe Nucleus That Express 5-HT3A Receptors Participate in Responses To Stress HormonesDocument9 pagesLi-2020-GABAergic Neurons in The Dorsal Raphe Nucleus That Express 5-HT3A Receptors Participate in Responses To Stress HormonesOmar Hernández GonzálezNo ratings yet
- SCNTFC RepDocument12 pagesSCNTFC RepRehaan FayazNo ratings yet
- HHS Public Access: Serotonin 2A Receptors Are A Stress Response System: Implications For Post-Traumatic Stress DisorderDocument22 pagesHHS Public Access: Serotonin 2A Receptors Are A Stress Response System: Implications For Post-Traumatic Stress DisorderDebbyNo ratings yet
- Serotonin Neurons, Neuroplasticity, and Homeostasis of Neural TissueDocument13 pagesSerotonin Neurons, Neuroplasticity, and Homeostasis of Neural TissueFuyumi RiaNo ratings yet
- Reviews: Thyroid Hormone Action and Brain DevelopmentDocument6 pagesReviews: Thyroid Hormone Action and Brain DevelopmentDewi PrasetiaNo ratings yet
- Neuro Nutrition 1983 v12n01 p038Document6 pagesNeuro Nutrition 1983 v12n01 p038LieMinkNo ratings yet
- Serotonin and MigraineDocument8 pagesSerotonin and MigrainexionimilliNo ratings yet
- Steroid Hormones and Their Action in Women's Brains: The Importance of Hormonal BalanceDocument15 pagesSteroid Hormones and Their Action in Women's Brains: The Importance of Hormonal BalanceBetão AviãoNo ratings yet
- Autacoid - SerotoninDocument4 pagesAutacoid - SerotoninJohn Ellard M. SaturnoNo ratings yet
- OpopDocument12 pagesOpopopopopopNo ratings yet
- Reseptor SerotoninDocument11 pagesReseptor SerotoninJuliana FeronNo ratings yet
- Beyond The Fixed Setpoint of The Hypothalamus-Pituitary-Thyroid AxisDocument12 pagesBeyond The Fixed Setpoint of The Hypothalamus-Pituitary-Thyroid AxisJuliany AscanioNo ratings yet
- Lessons in Exercise Neurobiology The Case of EndorphinsDocument6 pagesLessons in Exercise Neurobiology The Case of EndorphinsMarco NavaNo ratings yet
- Autism SpectrumDocument8 pagesAutism SpectrumNeutron ZionNo ratings yet
- INTRODUCTION TO NEUROPHARMACOLOGYyyDocument27 pagesINTRODUCTION TO NEUROPHARMACOLOGYyyEbad RazviNo ratings yet
- Etnografia 1Document9 pagesEtnografia 1gloriaNo ratings yet
- Yang Et Al, 2020 - O Papel Do BDNF Na Plasticidade Neural Na DepressãoDocument12 pagesYang Et Al, 2020 - O Papel Do BDNF Na Plasticidade Neural Na Depressãoprofessor do valeNo ratings yet
- Case Study - PTSD Final PaperDocument29 pagesCase Study - PTSD Final PaperMay PerezNo ratings yet
- Physiol 00027 2015Document17 pagesPhysiol 00027 2015Lilian EdeniNo ratings yet
- David Et Al 2009Document25 pagesDavid Et Al 2009izaquenNo ratings yet
- (14796805 - Journal of Endocrinology) The Role of Corticotropin-Releasing Factor in Depression and Anxiety DisordersDocument12 pages(14796805 - Journal of Endocrinology) The Role of Corticotropin-Releasing Factor in Depression and Anxiety Disorderskaren Daniela Puma FarinangoNo ratings yet
- Accorroni Et AlDocument17 pagesAccorroni Et Alp.sepulveda.fiNo ratings yet
- Etiology of Schizophrenia and Therapeutic Options: Gordana Rubeša, Lea Gudelj & Natalija KubinskaDocument8 pagesEtiology of Schizophrenia and Therapeutic Options: Gordana Rubeša, Lea Gudelj & Natalija KubinskaAwaluddin MisrangNo ratings yet
- Efectos Del Estrés Materno Prenatal Sobre La Serotonina y El Desarrollo FetalDocument6 pagesEfectos Del Estrés Materno Prenatal Sobre La Serotonina y El Desarrollo FetalkikemirandaNo ratings yet
- Canmedaj01342 0054Document4 pagesCanmedaj01342 0054joaoluizramoscetecNo ratings yet
- Oxytocin and Serotonin Brain Mechanisms in The Nonhuman PrimateDocument10 pagesOxytocin and Serotonin Brain Mechanisms in The Nonhuman PrimateidjacobsNo ratings yet
- Paper 23Document7 pagesPaper 23ramadhaniputrirezkiNo ratings yet
- Underwood 2018Document7 pagesUnderwood 2018Tristan TzaraNo ratings yet
- The Serotonergic System, The Pineal Gland & Side-EffectsDocument15 pagesThe Serotonergic System, The Pineal Gland & Side-EffectsElianKithaNo ratings yet
- ArtículoDocument12 pagesArtículoregina albertosNo ratings yet
- Mechanism of Action of LSDDocument10 pagesMechanism of Action of LSDgnuotakuNo ratings yet
- 1 s2.0 S0014488621001849 MainDocument14 pages1 s2.0 S0014488621001849 MainGregory AssonitisNo ratings yet
- Interface Between Hypothalamic-Pituitary-Adrenal Axis Andbrain-Derived Neurotrophic Factor in DepressionDocument13 pagesInterface Between Hypothalamic-Pituitary-Adrenal Axis Andbrain-Derived Neurotrophic Factor in DepressioncarlosNo ratings yet
- Effects of Serotonin Transporter Promoter Polymorphisms On Serotonin FunctionDocument9 pagesEffects of Serotonin Transporter Promoter Polymorphisms On Serotonin FunctionAnzari MuhammadNo ratings yet
- Tryptophan Metabolism and Gut-Brain HomeostasisDocument23 pagesTryptophan Metabolism and Gut-Brain HomeostasisLuísa OliveiraNo ratings yet
- Pet y SerotoninaDocument11 pagesPet y SerotoninaM.s. YamilethNo ratings yet
- Neurotransmitter and Behaviour: Serotonin and Anxiety: October 2011Document27 pagesNeurotransmitter and Behaviour: Serotonin and Anxiety: October 2011eva pravitasari nefertitiNo ratings yet
- Intro 1Document11 pagesIntro 1İlaydanur TaşdemirNo ratings yet
- REFERENCESDocument2 pagesREFERENCESİlaydanur TaşdemirNo ratings yet
- Debriefing Form TemplateDocument2 pagesDebriefing Form TemplateKelly AlevraNo ratings yet
- Informed Consent TemplateDocument3 pagesInformed Consent TemplateJan Diel100% (2)
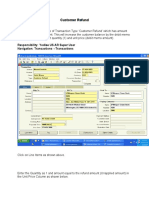
- Customer Refund: Responsibility: Yodlee US AR Super User Navigation: Transactions TransactionsDocument12 pagesCustomer Refund: Responsibility: Yodlee US AR Super User Navigation: Transactions TransactionsAziz KhanNo ratings yet
- Entity-Level Controls Fraud QuestionnaireDocument8 pagesEntity-Level Controls Fraud QuestionnaireKirby C. LoberizaNo ratings yet
- Air Augmented Rocket (285pages) Propulsion ConceptsDocument285 pagesAir Augmented Rocket (285pages) Propulsion ConceptsAlexandre PereiraNo ratings yet
- Biblehub Com Commentaries Matthew 3 17 HTMDocument21 pagesBiblehub Com Commentaries Matthew 3 17 HTMSorin TrimbitasNo ratings yet
- AmplifierDocument20 pagesAmplifierValerie StraussNo ratings yet
- The Machine StopsDocument14 pagesThe Machine StopsMICHAEL HARRIS USITANo ratings yet
- Advantages and Disadvantages of Global WarmingDocument11 pagesAdvantages and Disadvantages of Global WarmingA Aldika Farlis50% (2)
- 9.2 Volumetric Analysis PDFDocument24 pages9.2 Volumetric Analysis PDFJoaquinNo ratings yet
- Broshure JepanDocument6 pagesBroshure JepanIrwan Mohd YusofNo ratings yet
- ThesisDocument58 pagesThesisTirtha Roy BiswasNo ratings yet
- Argumentative Essay Project DescriptionDocument5 pagesArgumentative Essay Project DescriptionKaren Jh MoncayoNo ratings yet
- Fortune 2010Document14 pagesFortune 2010Stefan JenkinsNo ratings yet
- 7 - LESSON PLAN CULTURAL HERITAGE AND CULTURAL DIVERSITY - Lesson PlanDocument4 pages7 - LESSON PLAN CULTURAL HERITAGE AND CULTURAL DIVERSITY - Lesson PlanRute SobralNo ratings yet
- Module6 (Margie Ybanez)Document3 pagesModule6 (Margie Ybanez)Margie Ybañez82% (17)
- Public Administration 2000 - CSS ForumsDocument3 pagesPublic Administration 2000 - CSS ForumsMansoor Ali KhanNo ratings yet
- ACT December 2018 Form B05 PDFDocument54 pagesACT December 2018 Form B05 PDFPranav ChatiNo ratings yet
- Sample Internship PPTDocument19 pagesSample Internship PPTSangeeta JamadarNo ratings yet
- Behaviour of Investors in Indian Equity Markets: Submitted byDocument26 pagesBehaviour of Investors in Indian Equity Markets: Submitted byDibyanshu AmanNo ratings yet
- Moon and SaturnDocument4 pagesMoon and SaturnRamanasarmaNo ratings yet
- Unsaturated Polyester Resins: Influence of The Styrene Concentration On The Miscibility and Mechanical PropertiesDocument5 pagesUnsaturated Polyester Resins: Influence of The Styrene Concentration On The Miscibility and Mechanical PropertiesMamoon ShahidNo ratings yet
- Nurse-Patient Trust RelationshipDocument12 pagesNurse-Patient Trust RelationshipMarina Costa100% (1)
- Managing Ambiguity and ChangeDocument7 pagesManaging Ambiguity and ChangeTracey FeboNo ratings yet
- Class 12 Accountancy HHDocument58 pagesClass 12 Accountancy HHkomal barotNo ratings yet
- Analysis of Low-Frequency Passive Seismic Attributes in Maroun Oil Field, IranDocument16 pagesAnalysis of Low-Frequency Passive Seismic Attributes in Maroun Oil Field, IranFakhrur NoviantoNo ratings yet
- Full Download Social Animal 14th Edition Aronson Test BankDocument35 pagesFull Download Social Animal 14th Edition Aronson Test Banknaeensiyev100% (32)
- Internship Report On Effects of Promotion System On Employee Job Satisfaction of Janata Bank Ltd.Document57 pagesInternship Report On Effects of Promotion System On Employee Job Satisfaction of Janata Bank Ltd.Tareq Alam100% (1)
- Symbiosis National Aptitude Test (SNAP) 2004: InstructionsDocument21 pagesSymbiosis National Aptitude Test (SNAP) 2004: InstructionsHarsh JainNo ratings yet
- Impact of Micro FinanceDocument61 pagesImpact of Micro FinancePerry Arcilla SerapioNo ratings yet
- Asian Journal of ForestryDocument3 pagesAsian Journal of ForestryTeguh MuslimNo ratings yet
- India Marine Insurance Act 1963Document21 pagesIndia Marine Insurance Act 1963Aman GroverNo ratings yet
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (516)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (109)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (595)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)