You might also like
- The Brazilian Cerrado Vegetation and Threats To Its BiodiversityDocument8 pagesThe Brazilian Cerrado Vegetation and Threats To Its BiodiversityJackson RodriguesNo ratings yet
- CollembolaDocument13 pagesCollembolaMaría Mónica Bonilla PáezNo ratings yet
- Rocha 2004Document17 pagesRocha 2004Jaranna CoelhoNo ratings yet
- Nitrogen-Fixing Legume Tree Species For The Reclamation of Severely Degraded Lands in BrazilDocument11 pagesNitrogen-Fixing Legume Tree Species For The Reclamation of Severely Degraded Lands in BrazilFadhlan Husnul HafizhNo ratings yet
- A Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListDocument84 pagesA Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListJúnior SenseyNo ratings yet
- Janisovaal 2016 BullIAVSDocument17 pagesJanisovaal 2016 BullIAVSrangelbioNo ratings yet
- Rubiaceae Similarity and Conservation in Brazilian ForestsDocument13 pagesRubiaceae Similarity and Conservation in Brazilian ForestsJamesNo ratings yet
- Nitrogen and Phosphorus As Limiting Factors For Growth and Primary Production in A Ooded Savanna in The Venezuelan LlanosDocument11 pagesNitrogen and Phosphorus As Limiting Factors For Growth and Primary Production in A Ooded Savanna in The Venezuelan LlanosBichara DjimetNo ratings yet
- A Pollen Atlas of Premontane Woody and Herbaceous Communities From The Upland Savannas of Guayana, Venezuela - Leal Et Al. - 2011Document41 pagesA Pollen Atlas of Premontane Woody and Herbaceous Communities From The Upland Savannas of Guayana, Venezuela - Leal Et Al. - 2011vmsolartecNo ratings yet
- Flora and Life-Form Spectrum in An Area of Deciduous Thorn Woodland (Caatinga) in Northeastern, BrazilDocument11 pagesFlora and Life-Form Spectrum in An Area of Deciduous Thorn Woodland (Caatinga) in Northeastern, BrazilCecili MendesNo ratings yet
- DownloadDocument9 pagesDownloadpaolospadera72No ratings yet
- 2008 - Autotrophic Energy Sources For Paracheirodon Axelrodi Osteichthyes-Characidae in The Middle Negro River. Central Amazon, BrasilDocument9 pages2008 - Autotrophic Energy Sources For Paracheirodon Axelrodi Osteichthyes-Characidae in The Middle Negro River. Central Amazon, BrasilFabrício BarrosNo ratings yet
- 2007barbosaetal Biodivesidade-Conservao Lavrado RRDocument32 pages2007barbosaetal Biodivesidade-Conservao Lavrado RRmarianeNo ratings yet
- Relox and Camino Agusan Marsh PDFDocument10 pagesRelox and Camino Agusan Marsh PDFFritzie AtesNo ratings yet
- JACOMASSA, F. A. F. 2010. Espécies Arbóreas Nativas Da Mata Ciliar Da Bacia Hidrográfica Do Rio Lajeado Tunas, Na Região Do Alto Uruguai, RS PDFDocument6 pagesJACOMASSA, F. A. F. 2010. Espécies Arbóreas Nativas Da Mata Ciliar Da Bacia Hidrográfica Do Rio Lajeado Tunas, Na Região Do Alto Uruguai, RS PDFFábio André Facco JacomassaNo ratings yet
- Variación en La Composición de Comunidades de Aves en La Reserva de La Biosfera Montes Azules y Áreas Adyacentes, Chiapas, MéxicoDocument19 pagesVariación en La Composición de Comunidades de Aves en La Reserva de La Biosfera Montes Azules y Áreas Adyacentes, Chiapas, MéxicoFiorella Marycielo Cipriano RamosNo ratings yet
- Lee Et Al 2014 Diet and GeophagyDocument9 pagesLee Et Al 2014 Diet and GeophagyJim VillenaNo ratings yet
- Horvarth March Wolf 2001Document8 pagesHorvarth March Wolf 2001NachoMarchNo ratings yet
- Vansluys 2007Document7 pagesVansluys 2007samanthavieira.s39No ratings yet
- Caballero Et Al. 2019Document11 pagesCaballero Et Al. 2019ancginiNo ratings yet
- Boletn Unop Vol. 11 n2 2016 - GibbonsDocument22 pagesBoletn Unop Vol. 11 n2 2016 - GibbonsMario Tenorio MaldonadoNo ratings yet
- Ecology and conservation of the Red-tailed AmazonDocument16 pagesEcology and conservation of the Red-tailed AmazonjokercorvoNo ratings yet
- Diet Specialisation in BirdsDocument17 pagesDiet Specialisation in BirdsИванАнато́лийФранкоNo ratings yet
- Hernández Et Al., 2017. Diversidad y Patrones de Actividad de Mamíferos en Chiapas, MéxicoDocument13 pagesHernández Et Al., 2017. Diversidad y Patrones de Actividad de Mamíferos en Chiapas, MéxicoAlejandra HernándezNo ratings yet
- Dietz 1997Document17 pagesDietz 1997lassaad.el.hanafiNo ratings yet
- Wildediblefruits SimilipalDocument13 pagesWildediblefruits SimilipalAngel Eduardo Moreno AlboresNo ratings yet
- Agua ComposicionDocument10 pagesAgua Composicionnatu06lumarNo ratings yet
- Lopez Et Al, 2015 PDFDocument14 pagesLopez Et Al, 2015 PDFMario HerreraNo ratings yet
- art. Araujo et al. 2012. As aves e os ambientes em que elas ocorrem em umaDocument13 pagesart. Araujo et al. 2012. As aves e os ambientes em que elas ocorrem em umamateusfrancaNo ratings yet
- Lima Et Al. 2022 - NovaHedwigiaSemiaridchecklistDocument33 pagesLima Et Al. 2022 - NovaHedwigiaSemiaridchecklistJuliana LimaNo ratings yet
- Mendes Et Al. 2017 - Reproductive Phenology of Mauritia Flexuosa L. (Arecaceae)Document9 pagesMendes Et Al. 2017 - Reproductive Phenology of Mauritia Flexuosa L. (Arecaceae)Yakov Mario QuinterosNo ratings yet
- Impact of Exotic Pastures On Epigeic Arthropod Diversity and Contribution of Native and Exotic PlantDocument10 pagesImpact of Exotic Pastures On Epigeic Arthropod Diversity and Contribution of Native and Exotic PlantDomingas RosaNo ratings yet
- Crop Science - 2022 - Lara - Regrowth of Brachiariagrasses As Related To Stubble Characteristics Under N Fertilization andDocument11 pagesCrop Science - 2022 - Lara - Regrowth of Brachiariagrasses As Related To Stubble Characteristics Under N Fertilization andAryanda PrasetyaNo ratings yet
- CAP20ECOSISTEMASARAUCADocument32 pagesCAP20ECOSISTEMASARAUCADaniela VasquezNo ratings yet
- 24324-Texto Do Artigo-95554-1-10-20091119Document10 pages24324-Texto Do Artigo-95554-1-10-20091119Thiago de AlcântaraNo ratings yet
- RamosPereira Etal JAE 2009Document9 pagesRamosPereira Etal JAE 2009Joao ValsecchiNo ratings yet
- Diversity and Community Structure of Plants in Selected Areas Within Lake Mainit WatershedDocument19 pagesDiversity and Community Structure of Plants in Selected Areas Within Lake Mainit WatershedInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Mares ErnstDocument20 pagesMares ErnstJoana FerreiraNo ratings yet
- Check List: Serra Do Urubu, A Biodiversity Hot-Spot For Angiosperms in The Northern Atlantic Forest (Pernambuco, Brazil)Document25 pagesCheck List: Serra Do Urubu, A Biodiversity Hot-Spot For Angiosperms in The Northern Atlantic Forest (Pernambuco, Brazil)SôniaRodaNo ratings yet
- Saavedra-Rodríguez - Et - Al 2012Document9 pagesSaavedra-Rodríguez - Et - Al 2012Camilo HerreraNo ratings yet
- Attraction of Scarabaeinae (Coleoptera Scarabaeidae) To Different Baits in The Brazilian Amazon RegionDocument8 pagesAttraction of Scarabaeinae (Coleoptera Scarabaeidae) To Different Baits in The Brazilian Amazon RegionEduardo ValentimNo ratings yet
- Structure and diversity of three plant associations in the San Juan River DeltaDocument11 pagesStructure and diversity of three plant associations in the San Juan River DeltaCamil O'Hara MilloNo ratings yet
- 5 - Seasonal Variation in Leaf Traits Between Congeneric Savanna and Forest TreesDocument12 pages5 - Seasonal Variation in Leaf Traits Between Congeneric Savanna and Forest TreesAriadne Cristina De AntonioNo ratings yet
- Original Article /artículo Original: Héctor Aponte & D. Wilfredo RamírezDocument13 pagesOriginal Article /artículo Original: Héctor Aponte & D. Wilfredo RamírezKarla RicaldiiNo ratings yet
- CarvalhoeCarvalho 2012Document10 pagesCarvalhoeCarvalho 2012tmorato2No ratings yet
- Cochabamba Mountain-Finch breeding and calls documentedDocument9 pagesCochabamba Mountain-Finch breeding and calls documentedAndrea FuentesNo ratings yet
- Bonillaetal 2014Document15 pagesBonillaetal 2014biochoriNo ratings yet
- Guyana's Non-Timber Forest ProductsDocument350 pagesGuyana's Non-Timber Forest ProductsXenobia Delgado75% (4)
- Venado Cola BlancaDocument11 pagesVenado Cola BlancaErick Adrián Ramírez SantosNo ratings yet
- Forest Ecology and Management: A B A A C CDocument6 pagesForest Ecology and Management: A B A A C CJanaine RochaNo ratings yet
- Capitulo 11 PDFDocument26 pagesCapitulo 11 PDFAdilson SanchezNo ratings yet
- Bosques Secos en Ecuador y Su DiversidadDocument26 pagesBosques Secos en Ecuador y Su DiversidadJoseFrancisco MedinaNo ratings yet
- A Field Study of the Kansas Ant-Eating Frog, Gastrophryne olivaceaFrom EverandA Field Study of the Kansas Ant-Eating Frog, Gastrophryne olivaceaNo ratings yet
- Andradeetal2016 - Highland Grasslands at The Southern Tip of The Atlantic ForestDocument25 pagesAndradeetal2016 - Highland Grasslands at The Southern Tip of The Atlantic ForestKássia RamosNo ratings yet
- Gremios y Diversidad de Hormigas (Hymenoptera: Formicidae) en Tres Usos Del Suelo de Un Paisaje Cafetero Del Cauca-ColombiaDocument10 pagesGremios y Diversidad de Hormigas (Hymenoptera: Formicidae) en Tres Usos Del Suelo de Un Paisaje Cafetero Del Cauca-ColombiaAlex VillavicencioNo ratings yet
- A 26 V 60 N 2Document14 pagesA 26 V 60 N 2Diego Andres Moreno GaonaNo ratings yet
- Diversidad y Singularidad de La AvifaunaDocument16 pagesDiversidad y Singularidad de La Avifaunamyp20No ratings yet
- Density, Habitat Use, and Ranging Patterns of Red Howler Monkeys in A Colombian Andean ForestDocument10 pagesDensity, Habitat Use, and Ranging Patterns of Red Howler Monkeys in A Colombian Andean Forestt862092No ratings yet
- Castro & Kauffman 1998 Ecosystem Structure in The Brazilian Cerrado A Vegetation Gradient of AbovegroudDocument21 pagesCastro & Kauffman 1998 Ecosystem Structure in The Brazilian Cerrado A Vegetation Gradient of AbovegroudDeliane OliveiraNo ratings yet
- FrutalesdelaAmazonia-Isau 2008Document7 pagesFrutalesdelaAmazonia-Isau 2008Dario Santillan GarciaNo ratings yet
- The Forgotten PollinatorsDocument20 pagesThe Forgotten PollinatorsAntônio Gabriel Eleutério VianaNo ratings yet
- Conservation of Tree Species Through Sustainable Use How Can It Be Achieved in PracticeDocument11 pagesConservation of Tree Species Through Sustainable Use How Can It Be Achieved in PracticeppblancasNo ratings yet
- Puebla PrehistoryDocument39 pagesPuebla PrehistoryppblancasNo ratings yet
- Foraging and FarmingDocument17 pagesForaging and FarmingppblancasNo ratings yet
- Altieri Et Al., 1987Document10 pagesAltieri Et Al., 1987ppblancasNo ratings yet
- POH - PA44 Seminole G1000 NXiDocument388 pagesPOH - PA44 Seminole G1000 NXiroxyNo ratings yet
- West LP - M3 U6 L9Document10 pagesWest LP - M3 U6 L9Gretchen WestNo ratings yet
- Electrostatic Fields: ! R R R ! R Separation Distance (M) Between Q R /RDocument72 pagesElectrostatic Fields: ! R R R ! R Separation Distance (M) Between Q R /RRia Maria GeorgeNo ratings yet
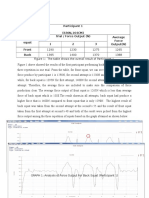
- Result: Participant 1 (630N, 164CM) Type of Squat Trial / Force Output (N) Average Force Output (N) 1 2 3 Front BackDocument5 pagesResult: Participant 1 (630N, 164CM) Type of Squat Trial / Force Output (N) Average Force Output (N) 1 2 3 Front BackpatenggNo ratings yet
- Sky TowerDocument19 pagesSky TowerAlcohol You LaterNo ratings yet
- Kinematic of MechanismDocument12 pagesKinematic of MechanismFennyFebrinaNurlitaSariNo ratings yet
- Raju OrdbmsDocument36 pagesRaju Ordbmssammy21791No ratings yet
- Mca 5th Sem AssignDocument16 pagesMca 5th Sem AssignVikas GuptaNo ratings yet
- Cisco ASR 9001 Router DatasheetDocument5 pagesCisco ASR 9001 Router DatasheetĐỗ TháiNo ratings yet
- User's Guide Materials Control: ProductionDocument52 pagesUser's Guide Materials Control: ProductionRanko LazeskiNo ratings yet
- Introduction To Structural AnalysisDocument15 pagesIntroduction To Structural AnalysisShereen RodrigoNo ratings yet
- Mini ProjectDocument45 pagesMini Projectmanohar manuNo ratings yet
- The Cost of Uncertainty For Nitrogen Fertilizer Management: A Sensitivity AnalysisDocument8 pagesThe Cost of Uncertainty For Nitrogen Fertilizer Management: A Sensitivity AnalysisSha-ReeNo ratings yet
- DOP-C02demo Exam Practice QuestionsDocument7 pagesDOP-C02demo Exam Practice Questionslovegeorge393No ratings yet
- Eseu MotivationalDocument6 pagesEseu MotivationalAlexandru RateaNo ratings yet
- Mechanical Engineering Laboratory InstrumentationDocument1 pageMechanical Engineering Laboratory InstrumentationGonzalo LNo ratings yet
- Drowsy Driving A Look at The Monitoring and Detection SystemDocument4 pagesDrowsy Driving A Look at The Monitoring and Detection SystemInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Econometric S Lecture 45Document31 pagesEconometric S Lecture 45Khurram AzizNo ratings yet
- Projet Hosseini2015Document11 pagesProjet Hosseini2015Høussàéîn Bèn MéssâõûdNo ratings yet
- Determination of Mechanical Properties For Cement-Treated Aggregate BaseDocument54 pagesDetermination of Mechanical Properties For Cement-Treated Aggregate BaseMarloNo ratings yet
- Analyzing Harmonic Polarities A Tonal NaDocument352 pagesAnalyzing Harmonic Polarities A Tonal Nagaia100% (1)
- BBS Model Question 2020Document44 pagesBBS Model Question 2020JALDIMAINo ratings yet
- CA - AssignmentReport - 2050020Document10 pagesCA - AssignmentReport - 2050020MINH NGUYỄN TÔNNo ratings yet
- Core Java - Munishwar GulatiDocument252 pagesCore Java - Munishwar Gulatimance1976No ratings yet
- Heat Exchanger BasicsDocument18 pagesHeat Exchanger BasicsJefferson WintersNo ratings yet
- Oxford Science 8Document258 pagesOxford Science 8Emily NuttingNo ratings yet
- Is Is at At: P,/P P, P,/PDocument4 pagesIs Is at At: P,/P P, P,/PNaqqash SajidNo ratings yet
- Tachyhydrite Camg CL 12H O: Crystal Data: Physical PropertiesDocument1 pageTachyhydrite Camg CL 12H O: Crystal Data: Physical Propertieskittipun khamprasoetNo ratings yet
- CB1 - Key Concepts in Biology (Paper 1 and 2)Document2 pagesCB1 - Key Concepts in Biology (Paper 1 and 2)Alhaji SowNo ratings yet
- Powder FlowDocument15 pagesPowder FlowSteven SastradiNo ratings yet