You might also like
- Lecture 3-Enzymes - KineticsDocument85 pagesLecture 3-Enzymes - Kineticslizzy.baongocNo ratings yet
- AT Unit 3-Part 1Document53 pagesAT Unit 3-Part 1ShivNo ratings yet
- Study On EnzymesDocument21 pagesStudy On EnzymesPragyan Kumar PradhanNo ratings yet
- Enzymes: Catalysts of Life's ReactionsDocument20 pagesEnzymes: Catalysts of Life's ReactionsEniola JayeolaNo ratings yet
- EnzymesDocument3 pagesEnzymesArabela SimanganNo ratings yet
- EnzymesDocument80 pagesEnzymesThisha MohanNo ratings yet
- Enzymes: Properties and KineticsDocument59 pagesEnzymes: Properties and Kineticsthamizh555100% (2)
- ENZYMES NOTES (Recovered)Document10 pagesENZYMES NOTES (Recovered)zipporahwaithera404No ratings yet
- 3 Enzymes PDFDocument50 pages3 Enzymes PDFBrianNo ratings yet
- Enzyme Kinetics Course Outcomes and TopicsDocument104 pagesEnzyme Kinetics Course Outcomes and Topicsclaudette_onaicna100% (2)
- Enzyme KineticsDocument14 pagesEnzyme Kineticsmradu1No ratings yet
- Final Study On EnzymeDocument9 pagesFinal Study On Enzymerosariopraveen007No ratings yet
- ERT211 CHP 1-2 PDFDocument27 pagesERT211 CHP 1-2 PDFsobiyamaragathavelNo ratings yet
- Sample 63Document9 pagesSample 63Rica NorcioNo ratings yet
- Enzyme Structure: OxidoreductasesDocument7 pagesEnzyme Structure: OxidoreductasesSamuel MutungaNo ratings yet
- 4 - EnzymesDocument31 pages4 - EnzymescheckmateNo ratings yet
- Biology ProjectDocument41 pagesBiology ProjectGanesan Siva67% (12)
- Enzyme Catalysis: Salahaddin-University-Erbil College of Education Chemistry DepartmentDocument11 pagesEnzyme Catalysis: Salahaddin-University-Erbil College of Education Chemistry DepartmentZana EdrisNo ratings yet
- Enzymes Basic Concepts and Kinetics 79Document66 pagesEnzymes Basic Concepts and Kinetics 79api-19824406No ratings yet
- 2 Bioprocess Engineering CH 2Document54 pages2 Bioprocess Engineering CH 2Fasil ManNo ratings yet
- GCE AS Biology 3 ENZYME NoteDocument11 pagesGCE AS Biology 3 ENZYME Notengnicole090607No ratings yet
- Enzymes ...Document13 pagesEnzymes ...Glen MangaliNo ratings yet
- 02 EnzymesDocument63 pages02 EnzymesFatish BanguraNo ratings yet
- Enzyme Catalysis: Mechanisms and Factors that Accelerate Reaction RatesDocument14 pagesEnzyme Catalysis: Mechanisms and Factors that Accelerate Reaction RatesAVNo ratings yet
- Enzymes Mbc213Document13 pagesEnzymes Mbc213prosperosamegieNo ratings yet
- Enzyme Kinetics NotesDocument2 pagesEnzyme Kinetics NotesMarc Imhotep Cray, M.D.No ratings yet
- PPT Biochemistry Chapter 2 (1)Document142 pagesPPT Biochemistry Chapter 2 (1)getasewalamir1No ratings yet
- Ma2003 Zainab Omolara Sanni - Practical Lesson 3Document6 pagesMa2003 Zainab Omolara Sanni - Practical Lesson 3geddy D.No ratings yet
- Enzyme Catalysis PDFDocument17 pagesEnzyme Catalysis PDFuzzal ahmedNo ratings yet
- Chapter 2: Diagnostic Enzymology: E+S Es E+PDocument87 pagesChapter 2: Diagnostic Enzymology: E+S Es E+PDereje KasuNo ratings yet
- Enzymes PDFDocument36 pagesEnzymes PDFAltamashNo ratings yet
- EnzymesDocument3 pagesEnzymesChampika NiranjaniNo ratings yet
- Lec 7 and 8 (Ch. 5) Enz KinDocument37 pagesLec 7 and 8 (Ch. 5) Enz KinRamy El-HadadNo ratings yet
- Enzymes - Michael L. ShulerDocument23 pagesEnzymes - Michael L. ShulerAndrésFelipeQuirogaMirandaNo ratings yet
- 06Enz1AMO SuarezDocument12 pages06Enz1AMO SuarezscasuarezNo ratings yet
- Nzymes: By: Mrs. Kalaivani Sathish. M. Pharm, Assistant Professor, Pims - PanipatDocument63 pagesNzymes: By: Mrs. Kalaivani Sathish. M. Pharm, Assistant Professor, Pims - Panipaturmila pandeyNo ratings yet
- Enzymology: Theresa May Chin TanDocument20 pagesEnzymology: Theresa May Chin TanVejerla PriyankaNo ratings yet
- Block 3BBCCT 107Document56 pagesBlock 3BBCCT 107robinNo ratings yet
- Enzymes: A Protein With Catalytic Properties Due To Its Power of Specific ActivationDocument35 pagesEnzymes: A Protein With Catalytic Properties Due To Its Power of Specific ActivationAkash SinghNo ratings yet
- Group Two'S Seminar Work: Topic: Enzyme Regulation Allosteric Regulation and Models OutlineDocument13 pagesGroup Two'S Seminar Work: Topic: Enzyme Regulation Allosteric Regulation and Models OutlineOluwasegun ModupeNo ratings yet
- Biochemistry Notes: Unit-5Th EnzymeDocument25 pagesBiochemistry Notes: Unit-5Th EnzymeKartik Rajpoot100% (1)
- Biochem ConceptsDocument9 pagesBiochem Conceptsdanena88No ratings yet
- Advanced Biochemistry EnzymesDocument108 pagesAdvanced Biochemistry EnzymesMalik TAHIRNo ratings yet
- 06Enz1AMO LICYAYODocument11 pages06Enz1AMO LICYAYOMohamidin MamalapatNo ratings yet
- 2 Biochemical Engineering CH 2Document54 pages2 Biochemical Engineering CH 2Fasil ManNo ratings yet
- Chapter 2: Diagnostic Enzymology: E+S Es E+PDocument72 pagesChapter 2: Diagnostic Enzymology: E+S Es E+PQananiisa Lammii Gammada100% (1)
- Enzyme TechnologyDocument19 pagesEnzyme TechnologySusmit NimjeNo ratings yet
- 01 Enzyme KineticsDocument5 pages01 Enzyme KineticsmwegiNo ratings yet
- Enzymes The Catalysts of LifeDocument43 pagesEnzymes The Catalysts of LifeKelsey WhitmoreNo ratings yet
- Enzymes: Structure, Function and Clinical ApplicationsDocument11 pagesEnzymes: Structure, Function and Clinical ApplicationsKathleen CedilloNo ratings yet
- c1.1 EnzymesDocument36 pagesc1.1 Enzymeshwqhd asjdhuaNo ratings yet
- 3 EnzymesDocument12 pages3 EnzymesmooshadabNo ratings yet
- EnzymesDocument46 pagesEnzymesShimmering MoonNo ratings yet
- General Principles of CatalysisDocument12 pagesGeneral Principles of CatalysisOLUWASEGUN K AfolabiNo ratings yet
- Enzymes Speed Up Chemical ReactionsDocument19 pagesEnzymes Speed Up Chemical ReactionsHanzala ShahidNo ratings yet
- Enzymes: Enzymes Homeostasis Activation EnergyDocument6 pagesEnzymes: Enzymes Homeostasis Activation EnergyGowthami MarreddyNo ratings yet
- Cell Bio Chapter 6Document32 pagesCell Bio Chapter 6GuteNo ratings yet
- Enzyme KineticsDocument13 pagesEnzyme KineticsKhushbu JainNo ratings yet
- Functional Metabolism: Regulation and AdaptationFrom EverandFunctional Metabolism: Regulation and AdaptationKenneth B. StoreyRating: 5 out of 5 stars5/5 (2)
- Glossary of Permafrost and Ground-Ice IPA 2005Document159 pagesGlossary of Permafrost and Ground-Ice IPA 2005NatitaGonzàlezDíazNo ratings yet
- Forcepoint Email Security Configuration Information v8.5Document64 pagesForcepoint Email Security Configuration Information v8.5ajay chaudharyNo ratings yet
- INTELLISPEC SERIE V-Páginas-31-46Document16 pagesINTELLISPEC SERIE V-Páginas-31-46Antonio Valencia VillejoNo ratings yet
- IJPC 13 4 Hormone Replacement TherapyDocument92 pagesIJPC 13 4 Hormone Replacement TherapyMatiasNo ratings yet
- Number SystemDocument4 pagesNumber SystemGlenn ThomasNo ratings yet
- Ultrasonic Couplant MSDSDocument2 pagesUltrasonic Couplant MSDSAnonymous uhqwokAeU0No ratings yet
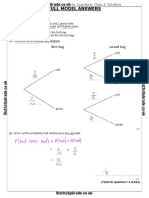
- Probability Tree Diagrams Solutions Mathsupgrade Co UkDocument10 pagesProbability Tree Diagrams Solutions Mathsupgrade Co UknatsNo ratings yet
- Calculating Maintenance and ReliabilityDocument7 pagesCalculating Maintenance and ReliabilityArdian P Noviatmoko100% (1)
- Science Grade 7Document8 pagesScience Grade 7Lacus ClyneNo ratings yet
- Taxation Management AssignmentDocument11 pagesTaxation Management AssignmentniraliNo ratings yet
- Breaugh Starke PDFDocument30 pagesBreaugh Starke PDFRichard YeongNo ratings yet
- Meta Hids by eBPFDocument27 pagesMeta Hids by eBPFninggou.yangNo ratings yet
- Research Trends On Environmental Energy and Vulnerability - 2021 - Energy andDocument27 pagesResearch Trends On Environmental Energy and Vulnerability - 2021 - Energy andCynthia Mac-beathNo ratings yet
- E-Way BillDocument1 pageE-Way BillShriyans DaftariNo ratings yet
- Rate of ChangeDocument22 pagesRate of ChangeTrisha MariehNo ratings yet
- SRV210 - 250TC Spare PartsDocument2 pagesSRV210 - 250TC Spare PartsEnzo SovittiNo ratings yet
- Filed & Entered: SBN 143271 SBN 165797 SBN 259014Document8 pagesFiled & Entered: SBN 143271 SBN 165797 SBN 259014Chapter 11 DocketsNo ratings yet
- SampleContract ShuttleDocument10 pagesSampleContract ShuttleLJ BNo ratings yet
- Pradeep Kumaresan Resume - Software Testing ExperienceDocument3 pagesPradeep Kumaresan Resume - Software Testing ExperienceSANTANo ratings yet
- Complete Tds CourseDocument32 pagesComplete Tds CourseAMLANNo ratings yet
- 3ms Squence 3 (Summary)Document3 pages3ms Squence 3 (Summary)toumi khadidjaNo ratings yet
- Video Conferencing Lesson PlanDocument6 pagesVideo Conferencing Lesson PlanANN JILLIAN JOYCE MONDONEDO100% (1)
- List of Trigonometric IdentitiesDocument16 pagesList of Trigonometric IdentitiesArnab NandiNo ratings yet
- v16 n3Document219 pagesv16 n3Mark ReinhardtNo ratings yet
- Business Letter WritingDocument13 pagesBusiness Letter WritingAlex Alexandru100% (1)
- 2020会社案内(英語版)Document18 pages2020会社案内(英語版)DAC ORGANIZERNo ratings yet
- Gray Iron Castings: Standard Specification ForDocument6 pagesGray Iron Castings: Standard Specification Forsafak kahramanNo ratings yet
- HTML MCQDocument22 pagesHTML MCQBuxar Students LifeNo ratings yet
- Photon BrochureDocument1 pagePhoton Brochureshwetha281889242No ratings yet
- AmboooDocument39 pagesAmboooTesfaye DegefaNo ratings yet