You might also like
- The Naturalist's Manual - Containing Descriptions of the Nests and Eggs of North American Birds (Turdidae - Tanagridae) also, Instructions for Collecting and Preserving Birds, Nests, Eggs and InsectsFrom EverandThe Naturalist's Manual - Containing Descriptions of the Nests and Eggs of North American Birds (Turdidae - Tanagridae) also, Instructions for Collecting and Preserving Birds, Nests, Eggs and InsectsNo ratings yet
- First Dinosaur Fossil DiscoveriesDocument4 pagesFirst Dinosaur Fossil DiscoveriesKimber LeeNo ratings yet
- Irish Elk - WikipediaDocument13 pagesIrish Elk - WikipediaDavid Gonzalez SotoNo ratings yet
- Dodo BonesDocument11 pagesDodo BonesFaisal MuhammadNo ratings yet
- 14 Fun Facts About Iguanodon: A 15-Minute BookFrom Everand14 Fun Facts About Iguanodon: A 15-Minute BookRating: 5 out of 5 stars5/5 (1)
- Acanthopholis: 1 HistoryDocument3 pagesAcanthopholis: 1 HistoryNupur PalNo ratings yet
- Baryonyx: Temporal Range: Early Cretaceous, 130-125 MaDocument16 pagesBaryonyx: Temporal Range: Early Cretaceous, 130-125 MaFelixNo ratings yet
- Rodrigues Starling: Jump To Navigation Jump To SearchDocument14 pagesRodrigues Starling: Jump To Navigation Jump To SearchTeodoro Miguel Carlos IsraelNo ratings yet
- 55Document2 pages55Mihailov VadimNo ratings yet
- British Birds - January 2021Document70 pagesBritish Birds - January 2021Dan BrownNo ratings yet
- Fauna Australia: 29. MacropodidaeDocument70 pagesFauna Australia: 29. MacropodidaeMuhammad SohailNo ratings yet
- Cryptozoology A To Z: The Encyclopedia Of Loch Monsters Sasquatch Chupacabras And Other Authentic MFrom EverandCryptozoology A To Z: The Encyclopedia Of Loch Monsters Sasquatch Chupacabras And Other Authentic MRating: 2.5 out of 5 stars2.5/5 (6)
- 100 Greatest DiscoveriesDocument2 pages100 Greatest DiscoveriesX GodinezNo ratings yet
- Did T. Rex Have Feathers?: Questions and Answers About DinosaursFrom EverandDid T. Rex Have Feathers?: Questions and Answers About DinosaursNo ratings yet
- Section 1: The Ielts HubDocument14 pagesSection 1: The Ielts HubTHE IELTS HUBNo ratings yet
- DINOSAURSDocument2 pagesDINOSAURSNhu Minh NguyenNo ratings yet
- Anomalocaris: Anomalocaris Anomalocaris ("Unlike Other Shrimp", or "Abnormal Shrimp") Is AnDocument9 pagesAnomalocaris: Anomalocaris Anomalocaris ("Unlike Other Shrimp", or "Abnormal Shrimp") Is Anenzo abrahamNo ratings yet
- 1111 Amazing Facts about Animals: Dinosaurs, dogs, lizards, insects, sharks, cats, birds, horses, snakes, spiders, fish and more!From Everand1111 Amazing Facts about Animals: Dinosaurs, dogs, lizards, insects, sharks, cats, birds, horses, snakes, spiders, fish and more!No ratings yet
- Natural History Museum Holotype of DynamosaurusDocument2 pagesNatural History Museum Holotype of Dynamosaurusdosonex274No ratings yet
- The Mistaken ExtinctionDocument30 pagesThe Mistaken Extinctionatulsahu24No ratings yet
- The Neanderthals: Who Were They and How did They LiveFrom EverandThe Neanderthals: Who Were They and How did They LiveRating: 5 out of 5 stars5/5 (1)
- Tyrannosauridae - WikipediaDocument16 pagesTyrannosauridae - WikipediaGopalNo ratings yet
- Neanderthal: 2 ClassificationDocument21 pagesNeanderthal: 2 ClassificationJohnnyBoz100% (1)
- Early HumansDocument26 pagesEarly HumansJohn DubzinskiNo ratings yet
- Piltdown ManDocument12 pagesPiltdown ManStephan WozniakNo ratings yet
- Darwin'S Voyage Around The WorldDocument4 pagesDarwin'S Voyage Around The WorldDavinci LegaspiNo ratings yet
- Koala - WikipediaDocument24 pagesKoala - WikipediaRico MartinezNo ratings yet
- How and Why Wonder Book of Extinct AnimalsDocument52 pagesHow and Why Wonder Book of Extinct Animalskett8233100% (5)
- Darwin at The Museum: of Species, Published in 1859Document3 pagesDarwin at The Museum: of Species, Published in 1859raj9792No ratings yet
- The Toad Stone A Rather Unlikely Gem PDFDocument3 pagesThe Toad Stone A Rather Unlikely Gem PDFDjefsky Krumluv100% (1)
- London S Science Musuem.Document26 pagesLondon S Science Musuem.MARLON ULISES MOROCHO ERREYESNo ratings yet
- Handouts For Evolution of SocietyDocument2 pagesHandouts For Evolution of SocietyJody CangrejoNo ratings yet
- Controversy The PlatypusDocument4 pagesControversy The Platypusskline3No ratings yet
- Moth Recorder Handbook Recommended Sources of InfoDocument8 pagesMoth Recorder Handbook Recommended Sources of InfoSergioOmarMachucaNo ratings yet
- AnkylosaurusDocument15 pagesAnkylosaurusShanmukha RongaliNo ratings yet
- 66Document2 pages66Mihailov VadimNo ratings yet
- Evolution Work Sheet - AnsweredDocument3 pagesEvolution Work Sheet - AnsweredJohn OsborneNo ratings yet
- Snake: From Wikipedia, The Free EncyclopediaDocument57 pagesSnake: From Wikipedia, The Free EncyclopediaVijay KaushalNo ratings yet
- John William DouglasDocument2 pagesJohn William DouglasFred PianistNo ratings yet
- Evoluton of Society Paleolithic and Mesolithic StageDocument1 pageEvoluton of Society Paleolithic and Mesolithic StageJody CangrejoNo ratings yet
- Istory of Jellyfish Envenomation: 3.1.1. Class Cubozoa - Cubozoan JellyfishDocument16 pagesIstory of Jellyfish Envenomation: 3.1.1. Class Cubozoa - Cubozoan JellyfishRangsiyo BeabeoNo ratings yet
- HR September 2020 Final 150dpi ZooViewDocument9 pagesHR September 2020 Final 150dpi ZooViewLuisNo ratings yet
- Brach IopodsDocument13 pagesBrach IopodsAthaariq AriqNo ratings yet
- Evolution - A Historical PerspectiveDocument13 pagesEvolution - A Historical PerspectiveRaphael MolanoNo ratings yet
- DINOSAURSDocument7 pagesDINOSAURSKhadija IjazNo ratings yet
- History of The Dodo HumeDocument25 pagesHistory of The Dodo HumeEuro VillaNo ratings yet
- Timeline of ExistenceDocument2 pagesTimeline of Existencecoffee creamNo ratings yet
- The History of 19th Cent.-Wps OfficeDocument12 pagesThe History of 19th Cent.-Wps OfficeAleck ZenonNo ratings yet
- Spotted Green PigeonDocument7 pagesSpotted Green PigeonVikas SainiNo ratings yet
- TiktaalikDocument14 pagesTiktaalikJupi THIRDNo ratings yet
- GomphotheriumDocument5 pagesGomphotheriumJupi THIRDNo ratings yet
- Estimate 2Document63 pagesEstimate 2Jupi THIRDNo ratings yet
- Joan RoofingplanDocument1 pageJoan RoofingplanJupi THIRDNo ratings yet
- Joan FloorplanDocument1 pageJoan FloorplanJupi THIRDNo ratings yet
- Bodega FloorplanDocument1 pageBodega FloorplanJupi THIRDNo ratings yet
- Bodega-Roof PlanDocument1 pageBodega-Roof PlanJupi THIRDNo ratings yet
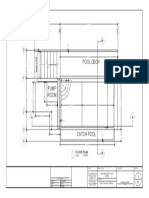
- Pool Deck: Floor PlanDocument1 pagePool Deck: Floor PlanJupi THIRDNo ratings yet
- Second Floor Plan: Bedroom-2 Bedroom-3Document1 pageSecond Floor Plan: Bedroom-2 Bedroom-3Jupi THIRDNo ratings yet
- 1.read and Match The Parts of The SentencesDocument2 pages1.read and Match The Parts of The SentencesнастяNo ratings yet
- Annie Ovenden Exibition 2017Document19 pagesAnnie Ovenden Exibition 2017Vitaliy ChuenkoNo ratings yet
- Compilation of Famous Architects and Their Architectures: Submitted By: Regine G. ChiongDocument13 pagesCompilation of Famous Architects and Their Architectures: Submitted By: Regine G. ChiongRegine G. ChiongNo ratings yet
- Roadmap Video The Roadmap ReportDocument2 pagesRoadmap Video The Roadmap ReportRodrigo Leao100% (2)
- КАРПЮК 5 Д Р Unit 7 1варDocument2 pagesКАРПЮК 5 Д Р Unit 7 1варЯна ГолобородькоNo ratings yet
- Sexual Dimorphism and Life History of Keichousaurus Hui (Reptilia: Sauropterygia)Document9 pagesSexual Dimorphism and Life History of Keichousaurus Hui (Reptilia: Sauropterygia)Adnana CebucNo ratings yet
- Meet AddyDocument79 pagesMeet AddykfontesNo ratings yet
- ROADMAP Report A2 Worksheet UnitDocument2 pagesROADMAP Report A2 Worksheet UnitAleNo ratings yet
- 2010 2011RDocument135 pages2010 2011RmsdoxseeNo ratings yet
- Session - 4Document9 pagesSession - 4CRISTIAN JOSEPH ARIAS LOPEZNo ratings yet
- DaldalDocument8 pagesDaldalabid aliNo ratings yet
- Host An Event - Ayala MuseumDocument10 pagesHost An Event - Ayala MuseumjizelhNo ratings yet
- Otto KunzliDocument2 pagesOtto Kunzlieileen gao100% (1)
- What Are The Etiquettes in A Museum and A GalleryDocument3 pagesWhat Are The Etiquettes in A Museum and A GalleryCARMELA MOSQUEDANo ratings yet
- SWOT Analysis For: Biomedical Museum, Kuala LumpurDocument2 pagesSWOT Analysis For: Biomedical Museum, Kuala Lumpurst nazirahNo ratings yet
- Moazzam Ali at ARTCITI GalleryDocument32 pagesMoazzam Ali at ARTCITI GalleryPrinceKhurramNo ratings yet
- Michelangelo and Anish KapoorDocument7 pagesMichelangelo and Anish Kapoorreemagotewala81No ratings yet
- Life in Color by Jacques Henri LartigueDocument1 pageLife in Color by Jacques Henri LartigueVallejoNo ratings yet
- Anthropology of SportDocument9 pagesAnthropology of SportAlfredo Arvin NievaNo ratings yet
- AAP101 - Unit I Lesson 1 Part 2 PDFDocument9 pagesAAP101 - Unit I Lesson 1 Part 2 PDFNizea Camille Navarro JavierNo ratings yet
- BululDocument2 pagesBululynnelNo ratings yet
- Curriculum Vitae (EN) - FARCAS PATRICIA ALEXA 3Document2 pagesCurriculum Vitae (EN) - FARCAS PATRICIA ALEXA 3Patricia Alexa FarcasNo ratings yet
- C V SEM Louis I. KahnDocument25 pagesC V SEM Louis I. KahnMina BcNo ratings yet
- Per Holck - "The Oseberg Ship Burial, Norway: New Thoughts On The Skeletons From The Grave Mound" (2006)Document27 pagesPer Holck - "The Oseberg Ship Burial, Norway: New Thoughts On The Skeletons From The Grave Mound" (2006)Before AfterNo ratings yet
- Forgery Beyond Deceit Fabrication Value and The Desire For Ancient Rome John North Hopkins Full ChapterDocument67 pagesForgery Beyond Deceit Fabrication Value and The Desire For Ancient Rome John North Hopkins Full Chapterchristopher.gomes879100% (6)
- Egypt's Amarna Letters Revealed Diplomacy in The Ancient WorldDocument6 pagesEgypt's Amarna Letters Revealed Diplomacy in The Ancient WorldMatheusFerreiraNo ratings yet
- CPAR 12-2ND Quarter Summative Test (2022)Document3 pagesCPAR 12-2ND Quarter Summative Test (2022)Ada Ricana100% (3)
- Archeology The Telltale ArtDocument6 pagesArcheology The Telltale Artapi-674602680No ratings yet
- Curtains Faucet Mirror Floor Lamp Closet: UNIT 4 TestDocument5 pagesCurtains Faucet Mirror Floor Lamp Closet: UNIT 4 TestNicole Montoya DelgadoNo ratings yet
- Riassunto DavidDocument2 pagesRiassunto Davidsofia canaleNo ratings yet
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseFrom EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseRating: 4.5 out of 5 stars4.5/5 (52)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (597)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (5)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (393)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (216)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (411)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (111)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)