You might also like
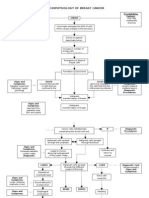
- Pathophysiology of Breast Cancer: Unkno Predisposing Factors: Precipitating FactorsDocument3 pagesPathophysiology of Breast Cancer: Unkno Predisposing Factors: Precipitating FactorsKevin Ercia100% (1)
- MCQs in PediatricsDocument4 pagesMCQs in Pediatricsarun babu93% (15)
- 1194-Article Text-5520-1-10-20200717Document8 pages1194-Article Text-5520-1-10-20200717Heryanti PusparisaNo ratings yet
- Resume VirologiDocument9 pagesResume Virologi752Puja Mardatula Aini D3 AkuntansiNo ratings yet
- Viruses 14 02073 PDFDocument17 pagesViruses 14 02073 PDFKRATIKA TYAGINo ratings yet
- Wijeratne2019 PDFDocument10 pagesWijeratne2019 PDFRudy Arindra WijayaNo ratings yet
- 2023 Article 378Document15 pages2023 Article 378Thaís ChouinNo ratings yet
- Dengue Virus ThesisDocument7 pagesDengue Virus Thesisafjvbpvce100% (1)
- Pone 0205248Document11 pagesPone 0205248Pawan MishraNo ratings yet
- JournalDocument12 pagesJournalDio SukardiNo ratings yet
- Molecular Technic of Virus IdentificationDocument13 pagesMolecular Technic of Virus Identificationpratiwi kusumaNo ratings yet
- Antigenic Evolution of Dengue Viruses Over 20 YearsDocument7 pagesAntigenic Evolution of Dengue Viruses Over 20 YearsAssao BachirNo ratings yet
- Journal PNTD 0001244Document10 pagesJournal PNTD 0001244Zulia Ahmad BurhaniNo ratings yet
- Laboratory Diagnosis of Dengue Infection: Current Techniques and Future StrategiesDocument9 pagesLaboratory Diagnosis of Dengue Infection: Current Techniques and Future StrategiesAndri YansyahNo ratings yet
- Dengue InglesDocument10 pagesDengue InglesalejandroNo ratings yet
- Dewi MegawatiDocument15 pagesDewi MegawatikasandraharahapNo ratings yet
- 24, Transmision en Defervescencia 2020Document5 pages24, Transmision en Defervescencia 2020Raquel FernandezNo ratings yet
- JJJJJJJJDocument4 pagesJJJJJJJJfreddyNo ratings yet
- 705 2019 Article 4266Document5 pages705 2019 Article 4266Melati CantikNo ratings yet
- Denv ThailandDocument16 pagesDenv ThailandGabriella StefanieNo ratings yet
- Denv MalaysiaDocument20 pagesDenv MalaysiaGabriella StefanieNo ratings yet
- Journal - Pone.0098785 1Document9 pagesJournal - Pone.0098785 1Haradhan PalNo ratings yet
- Journal Feng LiaoDocument19 pagesJournal Feng LiaoDewi KUNo ratings yet
- Dengue TOT Virology, Pathogenesis Lab Diag. 2023Document44 pagesDengue TOT Virology, Pathogenesis Lab Diag. 2023ATHIBAN VNo ratings yet
- 2015 Article 1022Document6 pages2015 Article 1022cindy clarissaNo ratings yet
- Dengue-Must KnowDocument16 pagesDengue-Must KnowJaylive AnziaNo ratings yet
- Performance of Simplexa Dengue Molecular Assay Compared To Conventional and SYBR Green RT-PCR For Detection of Dengue Infection in IndonesiaDocument10 pagesPerformance of Simplexa Dengue Molecular Assay Compared To Conventional and SYBR Green RT-PCR For Detection of Dengue Infection in IndonesiatyaminooNo ratings yet
- Current Understanding of The Pathogenesis of DenguDocument17 pagesCurrent Understanding of The Pathogenesis of DengucahyaNo ratings yet
- Vitamins and DengueDocument13 pagesVitamins and DengueShuvo DasNo ratings yet
- Immune-Mediated Cytokine Storm and Its Role in Severe DengueDocument12 pagesImmune-Mediated Cytokine Storm and Its Role in Severe DengueAkila SwaminathanNo ratings yet
- Long-Term Safety and Immunogenicity of A Tetravalent May 2022Document8 pagesLong-Term Safety and Immunogenicity of A Tetravalent May 2022Reinaldi octaNo ratings yet
- 1 s2.0 S1876034123003143 MainDocument5 pages1 s2.0 S1876034123003143 MainThaís ChouinNo ratings yet
- The Future of Dengue Vaccines - LancetDocument3 pagesThe Future of Dengue Vaccines - LancetnabinshresthacyzNo ratings yet
- Largest Dengue Outbreak (2022) in Nepal: Original Article Nepal Med Coll J March 2023 25 (1) : 73-8Document6 pagesLargest Dengue Outbreak (2022) in Nepal: Original Article Nepal Med Coll J March 2023 25 (1) : 73-8hunzala shamirNo ratings yet
- DengueDocument32 pagesDenguetummalapalli venkateswara rao100% (3)
- 529 With Cover Page v2Document8 pages529 With Cover Page v2Dwita Ayu IsrawatiNo ratings yet
- Tropmed 83 690 PDFDocument6 pagesTropmed 83 690 PDFabbhyasa5206No ratings yet
- 1.1 The DiseaseDocument33 pages1.1 The DiseaseabdulNo ratings yet
- SinglaDocument25 pagesSinglakasandraharahapNo ratings yet
- Dengue Virus Infection and Neurological ManifestationsDocument9 pagesDengue Virus Infection and Neurological Manifestationsjtd9h2xp5hNo ratings yet
- Review: Dengue ImmunopathologyDocument8 pagesReview: Dengue ImmunopathologyKhalisah dinahNo ratings yet
- Suppression of Virus Specific Immune Responses by IL-10 in Acute Dengue InfectionDocument10 pagesSuppression of Virus Specific Immune Responses by IL-10 in Acute Dengue InfectionThuongNguyen1981No ratings yet
- Emergence of Dengue Virus Type 4 (Genotype I) in IndiaDocument4 pagesEmergence of Dengue Virus Type 4 (Genotype I) in IndiajudodododoNo ratings yet
- Sero-Epidemiology of Dengue Virus Infection in Cities of IndonesiaDocument4 pagesSero-Epidemiology of Dengue Virus Infection in Cities of IndonesiaKris Cahyo MulyatnoNo ratings yet
- Jose Research v2Document10 pagesJose Research v2Jose Emmanuel FranciaNo ratings yet
- Severe Dengue in The Intensive Care UnitDocument18 pagesSevere Dengue in The Intensive Care UnitHitallo BezerraNo ratings yet
- TLR2 Axis On Peripheral Blood Mononuclear Cells Regulates Inflammatory Responses To Circulating, Non-Infectious Immature Dengue Virus ParticlesDocument37 pagesTLR2 Axis On Peripheral Blood Mononuclear Cells Regulates Inflammatory Responses To Circulating, Non-Infectious Immature Dengue Virus ParticlesS. ChenNo ratings yet
- Dengue Infection Global Importance, Immunopathology and ManagementDocument5 pagesDengue Infection Global Importance, Immunopathology and ManagementAlice in wonderlandNo ratings yet
- DengueDocument4 pagesDengueredyhataNo ratings yet
- Respon Imun MersDocument7 pagesRespon Imun MersKezia MarsilinaNo ratings yet
- Typing of Dengue Viruses in Clinical Specimens and Mosquitoes by Single-Tube Multiplex Reverse Transcriptase PCRDocument6 pagesTyping of Dengue Viruses in Clinical Specimens and Mosquitoes by Single-Tube Multiplex Reverse Transcriptase PCRaracnidoNo ratings yet
- O Dengue 2articulaDocument12 pagesO Dengue 2articulavillanueva23749No ratings yet
- 2016 Article 1849 PDFDocument8 pages2016 Article 1849 PDFdydy_7193No ratings yet
- 2016 Article 1668Document7 pages2016 Article 1668Rahmiati LaoNo ratings yet
- Biosensors and Bioelectronics: Daniel Wasik, Ashok Mulchandani, Marylynn V. YatesDocument6 pagesBiosensors and Bioelectronics: Daniel Wasik, Ashok Mulchandani, Marylynn V. YatesSamar D'melloNo ratings yet
- 01 Daljeet KaurDocument9 pages01 Daljeet KaurOcta VianiNo ratings yet
- Estudio 2 PDFDocument28 pagesEstudio 2 PDFCr A AleydaNo ratings yet
- Dengue 1Document14 pagesDengue 1FikaAuliaNo ratings yet
- ID Sebaran Serotipe Virus Dengue Di PontianDocument6 pagesID Sebaran Serotipe Virus Dengue Di PontianHera sasmitaNo ratings yet
- DengueDocument30 pagesDengueSanjeevNo ratings yet
- 32 165 1 PB PDFDocument5 pages32 165 1 PB PDFKennyShelva266No ratings yet
- 2022 - Fluid Management in Dengue Critical PhaseDocument7 pages2022 - Fluid Management in Dengue Critical PhasegiseladlrNo ratings yet
- Arterial DiseaseDocument191 pagesArterial DiseaseAura DiscyacittaNo ratings yet
- Circulationaha 118 030099Document19 pagesCirculationaha 118 030099Aura DiscyacittaNo ratings yet
- Steroid Induced Diabetes and Hyperglycaemia Part 2 ManagementDocument5 pagesSteroid Induced Diabetes and Hyperglycaemia Part 2 ManagementAura DiscyacittaNo ratings yet
- Practical Diabetes International - November December 1996 - Dunning - Corticosteroid Medications and Diabetes MellitusDocument3 pagesPractical Diabetes International - November December 1996 - Dunning - Corticosteroid Medications and Diabetes MellitusAura DiscyacittaNo ratings yet
- Mayer2013 Genetic Polymorphisms Associated With Intervertebral Disc DegenerationDocument19 pagesMayer2013 Genetic Polymorphisms Associated With Intervertebral Disc DegenerationAura DiscyacittaNo ratings yet
- Morris 2020 The Role of The Locus Coeruleus in TheDocument18 pagesMorris 2020 The Role of The Locus Coeruleus in TheAura DiscyacittaNo ratings yet
- Marsland 2017 The Effects of Acute Psychological Stress On Circulating and Stimulated Inflammatory Markers A Systematic Review and Meta AnalysisDocument41 pagesMarsland 2017 The Effects of Acute Psychological Stress On Circulating and Stimulated Inflammatory Markers A Systematic Review and Meta AnalysisAura DiscyacittaNo ratings yet
- Marcinkiewcz2016 Serotonin Engages An Anxiety and Fear-Promoting Circuit in AmygdalaDocument19 pagesMarcinkiewcz2016 Serotonin Engages An Anxiety and Fear-Promoting Circuit in AmygdalaAura DiscyacittaNo ratings yet
- GIGANTISMDocument26 pagesGIGANTISMAura DiscyacittaNo ratings yet
- Hookworm Infestation Causes Iron Deficiency Anemia: October 2017Document2 pagesHookworm Infestation Causes Iron Deficiency Anemia: October 2017Aura DiscyacittaNo ratings yet
- Case 3Document16 pagesCase 3Aura DiscyacittaNo ratings yet
- Endocrine SystemDocument2 pagesEndocrine SystemAura DiscyacittaNo ratings yet
- Ekg For The BeginnersDocument59 pagesEkg For The BeginnersAura DiscyacittaNo ratings yet
- Tugas Tutorial 2-Aura Discyacitta 1710211036 Kelas Tutorial C2Document2 pagesTugas Tutorial 2-Aura Discyacitta 1710211036 Kelas Tutorial C2Aura DiscyacittaNo ratings yet
- Medical Self Declaration FormDocument1 pageMedical Self Declaration FormVarun SinglaNo ratings yet
- 2.tukak PeptikDocument42 pages2.tukak PeptikEfvi VhyLiaNo ratings yet
- Peripheral Nerve Field Stimulation For Sacroiliac Joint PainDocument4 pagesPeripheral Nerve Field Stimulation For Sacroiliac Joint PainAna Cristina MartinsNo ratings yet
- Peme Reme Form ADocument1 pagePeme Reme Form AHector ValleNo ratings yet
- "Bloody Scours": Swine DysenteryDocument29 pages"Bloody Scours": Swine DysenteryleaNo ratings yet
- Manejo de Hipermenorrea en AdolescentesDocument9 pagesManejo de Hipermenorrea en AdolescentesDavid VargasNo ratings yet
- Hospital and Industrial Hygiene Part 1 N.V (2 Files Merged)Document21 pagesHospital and Industrial Hygiene Part 1 N.V (2 Files Merged)giNo ratings yet
- Medication: Expected Pharmacological Action Therapeutic UseDocument1 pageMedication: Expected Pharmacological Action Therapeutic UseMike EveretteNo ratings yet
- Peppermint LetterDocument2 pagesPeppermint LetterKelli AdamsNo ratings yet
- Blumenfeld Neuroanatomy Ch. 5 SummaryDocument5 pagesBlumenfeld Neuroanatomy Ch. 5 SummaryMeeraNo ratings yet
- Why Does My Patient Limphadenopathy or Splenomegaly?Document14 pagesWhy Does My Patient Limphadenopathy or Splenomegaly?lacmftcNo ratings yet
- Intestinal ObstructionDocument59 pagesIntestinal ObstructionGeorge GuruNo ratings yet
- Early Warning SystemDocument6 pagesEarly Warning SystemElizeus HaninditoNo ratings yet
- Nosocomial Infections: Causes, Controls, Preventions and Future ChallengesDocument11 pagesNosocomial Infections: Causes, Controls, Preventions and Future ChallengesAy GlassesNo ratings yet
- Knowledge and Attitude Towards Malaria Knowlesi: A Case Study at Senaning Village, Ketungau Hulu Sub-District Sintang DistrictDocument15 pagesKnowledge and Attitude Towards Malaria Knowlesi: A Case Study at Senaning Village, Ketungau Hulu Sub-District Sintang DistrictAbdul RamalangiNo ratings yet
- CDC Welcome LetterDocument5 pagesCDC Welcome Letterapi-242666861No ratings yet
- Parenteral Therapy:: Intravenous Therapy (IVT) or VenipunctureDocument3 pagesParenteral Therapy:: Intravenous Therapy (IVT) or VenipunctureMabes100% (1)
- Evaluation of Microscopic HematuriaDocument15 pagesEvaluation of Microscopic HematuriaAndi SusiloNo ratings yet
- GDS K-6 - Newborn Physical ExaminationDocument44 pagesGDS K-6 - Newborn Physical ExaminationJosephine IrenaNo ratings yet
- Uk Fast ForwardDocument19 pagesUk Fast Forwardapi-549064749No ratings yet
- OsteoarthritisDocument11 pagesOsteoarthritisadddNo ratings yet
- Stago Vol 5 Issue 2 2015Document10 pagesStago Vol 5 Issue 2 2015PABLO ZAMUDIONo ratings yet
- Too Tired EDS and Fatigue - Brad TinkleDocument34 pagesToo Tired EDS and Fatigue - Brad TinkleGèniaNo ratings yet
- Review of Medical Microbiology and Immunology (PDFDrive) - Pages-322-343,365-371Document29 pagesReview of Medical Microbiology and Immunology (PDFDrive) - Pages-322-343,365-371Mae Rose Charlene MendozaNo ratings yet
- First Conditional: "If You Fall, I Will Be There." - The FloorDocument3 pagesFirst Conditional: "If You Fall, I Will Be There." - The FloorgastonNo ratings yet
- Diagnostic Skills of Acupuncture: Lecture By: M.K. SastryDocument106 pagesDiagnostic Skills of Acupuncture: Lecture By: M.K. SastryLupa IngatanNo ratings yet
- Atelektasis Jurnal RadiologiDocument19 pagesAtelektasis Jurnal RadiologiRachmi MerrinaNo ratings yet