You might also like
- Identification of Pathogenic Bacteria in Clinical Microbiology LaboratoryDocument70 pagesIdentification of Pathogenic Bacteria in Clinical Microbiology Laboratoryamare100% (4)
- Sources of Epidemiological DataDocument3 pagesSources of Epidemiological Datageorgeloto1283% (6)
- Pnas01519 0334Document6 pagesPnas01519 0334medtubeburnerNo ratings yet
- Ozolione Fmolb 2020-Excessive Promoters As Silencers of Genes Horizontally Acquired by Escherichia ColiDocument9 pagesOzolione Fmolb 2020-Excessive Promoters As Silencers of Genes Horizontally Acquired by Escherichia ColiDiegoNo ratings yet
- DNA Repair Protein Involved in Heart and Blood DevelopmentDocument12 pagesDNA Repair Protein Involved in Heart and Blood DevelopmentSol Jumaide WerbleNo ratings yet
- Vasquezii FVIDocument9 pagesVasquezii FVInfernandezNo ratings yet
- Biochem Markers.2006Document16 pagesBiochem Markers.2006Wahyu W'cas PamungkasNo ratings yet
- Url-2003 Parvovirus en Neuronas de GatoDocument5 pagesUrl-2003 Parvovirus en Neuronas de GatocristianNo ratings yet
- Occurrence of Honeysuckle Yellow Vein Virus (HYVV) Containing A Monopartite DNA-A Genome in KoreaDocument10 pagesOccurrence of Honeysuckle Yellow Vein Virus (HYVV) Containing A Monopartite DNA-A Genome in KoreaFrontiersNo ratings yet
- AssstDocument5 pagesAssstDiegoNo ratings yet
- Gren 2016Document38 pagesGren 2016wildan ariefNo ratings yet
- Non Pumping Function of The PumpDocument9 pagesNon Pumping Function of The PumpnkmwisNo ratings yet
- Delalande1998 PDFDocument3 pagesDelalande1998 PDFNada ZnaidiNo ratings yet
- Farkas Parvo 2004Document7 pagesFarkas Parvo 2004Sandra EßbauerNo ratings yet
- First Report of Peanut Stunt Virus On Glycine MaxDocument2 pagesFirst Report of Peanut Stunt Virus On Glycine MaxemerensianaNo ratings yet
- Parakua Acara 4 InggrisDocument23 pagesParakua Acara 4 InggrisPuri RahmaNo ratings yet
- tmpD473 TMPDocument14 pagestmpD473 TMPFrontiersNo ratings yet
- 1 s2.0 S0006349518316114 MainDocument1 page1 s2.0 S0006349518316114 MainDiego TulcanNo ratings yet
- Ijms 20 00107Document20 pagesIjms 20 00107SRI RAHMAYUNINo ratings yet
- Fendo 13 1005863Document21 pagesFendo 13 1005863Wahyu PamungkasNo ratings yet
- 2019 Article 421DTNDocument8 pages2019 Article 421DTNDulce Maria AngelesNo ratings yet
- 03 Zwiewka Et Al. 2011 Cell ResDocument12 pages03 Zwiewka Et Al. 2011 Cell ResMugurel I. FeraruNo ratings yet
- First Report of Amblyomma Sp. Collected From Varanus SalvatorDocument6 pagesFirst Report of Amblyomma Sp. Collected From Varanus Salvatorbambang awanNo ratings yet
- Assembly of Protein Complexes Restricts DiffusionDocument15 pagesAssembly of Protein Complexes Restricts DiffusionRifqi Fathul ArroisiNo ratings yet
- Tumor Suppressor APC Blocks DNA Polymerase - Dependent Strand Displacement Synthesis During Long Patch But Not Short Patch Base Excision Repair and Increases Sensitivity To Methylmethane SulfonateDocument8 pagesTumor Suppressor APC Blocks DNA Polymerase - Dependent Strand Displacement Synthesis During Long Patch But Not Short Patch Base Excision Repair and Increases Sensitivity To Methylmethane SulfonateBiosynthesisNo ratings yet
- 2003-Culliman-PERK and NRF2Document12 pages2003-Culliman-PERK and NRF2HaiNo ratings yet
- Mizgalska Potempa Et-Al Genome Sequence of Porphyromonas Gingivalis 2014Document2 pagesMizgalska Potempa Et-Al Genome Sequence of Porphyromonas Gingivalis 2014Nathaly Andrea Delgadillo SalgadoNo ratings yet
- 2008 - Whole Genome PlatypusDocument11 pages2008 - Whole Genome Platypusj_wingerdNo ratings yet
- Benh VirusDocument7 pagesBenh VirusThu Hien NguyenNo ratings yet
- Journal of Life Sciences & BiomedicineDocument7 pagesJournal of Life Sciences & BiomedicineBiomedicince journalNo ratings yet
- TMP F054Document3 pagesTMP F054FrontiersNo ratings yet
- Bi 0274322Document10 pagesBi 0274322Carlos MeirellesNo ratings yet
- Rapid de Novo Assembly of The European Eel Genome From Nanopore Sequencing ReadsDocument13 pagesRapid de Novo Assembly of The European Eel Genome From Nanopore Sequencing ReadsMuhammad SiddiqNo ratings yet
- Tsuda2012 P AlviolaDocument7 pagesTsuda2012 P AlviolajadeNo ratings yet
- Cloning, Expression and Purification of Mouse Vegf (Vascular Endothelial Growth Factor) in E. Coli 01 FebDocument4 pagesCloning, Expression and Purification of Mouse Vegf (Vascular Endothelial Growth Factor) in E. Coli 01 FebGJESRNo ratings yet
- Eric 1779Document5 pagesEric 1779Daniela Araya GonzalezNo ratings yet
- Modelo de ObesidadDocument8 pagesModelo de ObesidadJorge Hantar Touma LazoNo ratings yet
- Electrical Characters On Surface of Apoptotic Cell in Maize Roots Induced by CytotoxinsDocument10 pagesElectrical Characters On Surface of Apoptotic Cell in Maize Roots Induced by CytotoxinsUmiatin RamdhaniNo ratings yet
- Xu2015 Article RapidGenerationAndAnalysisOfExDocument13 pagesXu2015 Article RapidGenerationAndAnalysisOfExIrsyadKamalNo ratings yet
- Comparing Two Intestinal Porcine Epithelial Cell Lines (Ipecs) : Morphological Differentiation, Function and MetabolismDocument20 pagesComparing Two Intestinal Porcine Epithelial Cell Lines (Ipecs) : Morphological Differentiation, Function and MetabolismCristina BulgaruNo ratings yet
- Hippo Signaling in Epithelial Stem Cells: ReviewDocument7 pagesHippo Signaling in Epithelial Stem Cells: ReviewAlfi SyahriNo ratings yet
- Adaptive Evolution and Functional Divergence of Pepsin Gene FamilyDocument10 pagesAdaptive Evolution and Functional Divergence of Pepsin Gene FamilyharisankarhsNo ratings yet
- Mass-spectrometric and bioinformatic analysis of eEF1Bγ interactome in the cytoplasmic fraction of A549 cellsDocument11 pagesMass-spectrometric and bioinformatic analysis of eEF1Bγ interactome in the cytoplasmic fraction of A549 cellsАнна ШаповаловаNo ratings yet
- Cladistic Relationships Among The Pleurotus Ostreatus Complex, The Pleurotus Pulmonarius Complex, and Pleurotus Eryngii Based On The Mitochondrial Small Subunit Ribosomal DNA Sequence AnalysisDocument6 pagesCladistic Relationships Among The Pleurotus Ostreatus Complex, The Pleurotus Pulmonarius Complex, and Pleurotus Eryngii Based On The Mitochondrial Small Subunit Ribosomal DNA Sequence AnalysisAlejandro Murillo V.No ratings yet
- Ecotoxicology and Environmental Safety: Tong Liu, Luqing Pan, Qian Jin, Yuefeng CaiDocument11 pagesEcotoxicology and Environmental Safety: Tong Liu, Luqing Pan, Qian Jin, Yuefeng CaiArdy SetiaNo ratings yet
- Early Sporulation GeneDocument5 pagesEarly Sporulation GeneStephen G. SabinayNo ratings yet
- Journal Pone 0247180Document20 pagesJournal Pone 0247180Rahul ShelkeNo ratings yet
- Immunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient MouseDocument5 pagesImmunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient Mouselily rogaNo ratings yet
- Appl. Environ. Microbiol.-2005-Barak-5685-91Document7 pagesAppl. Environ. Microbiol.-2005-Barak-5685-91NORMA SABIONo ratings yet
- SSR130531 Wyz (1) .Doc CorrectedDocument7 pagesSSR130531 Wyz (1) .Doc CorrectedjosekinNo ratings yet
- Pan Et Al 2019Document8 pagesPan Et Al 2019misilpiyapiyoNo ratings yet
- 1 s2.0 S0093691X19303851 MainDocument14 pages1 s2.0 S0093691X19303851 Mainyujiao MaNo ratings yet
- Pahala 1Document5 pagesPahala 1Daeng MangkaNo ratings yet
- AcidoDocument6 pagesAcidoDark WolfNo ratings yet
- MitokondriaDocument9 pagesMitokondriaMakinrame MakinmurahNo ratings yet
- Actina 1Document2 pagesActina 1Anonymous yeHhCYNo ratings yet
- The Role of WNT Signaling in Axis Development and Stem Cell Regulation in CnidariansDocument6 pagesThe Role of WNT Signaling in Axis Development and Stem Cell Regulation in CnidariansEliasNo ratings yet
- The Unfolded Protein Response Is Triggered by A Plant Viral Movement ProteinDocument15 pagesThe Unfolded Protein Response Is Triggered by A Plant Viral Movement ProteinFrontiersNo ratings yet
- Bahan Paper PatoDocument6 pagesBahan Paper PatoKhairul Ihsan Pengusaha MudaNo ratings yet
- Animal Reproduction Science: Characterization of Prostaglandin E Receptors (EP2, EP4) in The Horse OviductDocument7 pagesAnimal Reproduction Science: Characterization of Prostaglandin E Receptors (EP2, EP4) in The Horse OviductDiana RNo ratings yet
- Chen 2007Document8 pagesChen 2007yuliatyNo ratings yet
- DNA - Recombination Interactions and Repair: Proceedings of the FEBS Symposium on DNA, Liblice, 1979From EverandDNA - Recombination Interactions and Repair: Proceedings of the FEBS Symposium on DNA, Liblice, 1979S. ZadrazilNo ratings yet
- Guidence BookletDocument63 pagesGuidence BookletSudeepta SinghNo ratings yet
- Jumbophages - Overview - Review ArticleDocument9 pagesJumbophages - Overview - Review ArticleSudeepta SinghNo ratings yet
- Viruses 13 00063 v3Document42 pagesViruses 13 00063 v3Sudeepta SinghNo ratings yet
- Lysin - Fmicb 13 856473Document11 pagesLysin - Fmicb 13 856473Sudeepta SinghNo ratings yet
- Lysin - Comp Rev Food Sci Food Safe - 2023 - Kocot - Phages and Engineered Lysins As An Effective Tool To Combat Gram NegativeDocument32 pagesLysin - Comp Rev Food Sci Food Safe - 2023 - Kocot - Phages and Engineered Lysins As An Effective Tool To Combat Gram NegativeSudeepta SinghNo ratings yet
- 1ST Lab MidtermDocument4 pages1ST Lab MidtermMary Jane TiangsonNo ratings yet
- Syndromic Management of Sexually Transmitable Infections Reference Manual (2) - 1Document116 pagesSyndromic Management of Sexually Transmitable Infections Reference Manual (2) - 1Keneni100% (1)
- COVID19 by DR CL GuruduttDocument2 pagesCOVID19 by DR CL GuruduttkarthikNo ratings yet
- Jurnal 2Document3 pagesJurnal 2danna aflahNo ratings yet
- Tinea Cruris Abq JournalDocument3 pagesTinea Cruris Abq JournalGustiandari FidhyaNo ratings yet
- Solution Manual For Generalized Linear Models 0205377939Document24 pagesSolution Manual For Generalized Linear Models 0205377939BrianGarzapeyn100% (35)
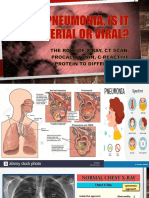
- Pneumonia, Is It Bacterial or Viral FINAL EDITINGDocument31 pagesPneumonia, Is It Bacterial or Viral FINAL EDITINGdv prnt1909No ratings yet
- F Y) Y, 0 y 3: Fuad Adrian Iskandar 1606843722 PR-I Matematika Terapan (Statistik)Document2 pagesF Y) Y, 0 y 3: Fuad Adrian Iskandar 1606843722 PR-I Matematika Terapan (Statistik)Nadia KarimaNo ratings yet
- Universal Precautions PPT FinalDocument50 pagesUniversal Precautions PPT Finalmarthinfp100% (1)
- CLINICAL PLACEMENT PERMIT CHECKLIST Brock UniversityDocument2 pagesCLINICAL PLACEMENT PERMIT CHECKLIST Brock UniversityroldinpgNo ratings yet
- Aniwada 1332015 BJMMR23249 DSNDocument11 pagesAniwada 1332015 BJMMR23249 DSNelias aniwadaNo ratings yet
- Grade 10 Reproductive HealthDocument26 pagesGrade 10 Reproductive HealthMissy BanguisNo ratings yet
- Negative: What Does It Mean To Have A Test Result?Document2 pagesNegative: What Does It Mean To Have A Test Result?Seiyi KohayagawaNo ratings yet
- The Black DeathDocument6 pagesThe Black DeathjaniNo ratings yet
- White Blood CellsDocument20 pagesWhite Blood CellsHarram SajjadNo ratings yet
- AF SeedingDocument6 pagesAF SeedingdonkeyendutNo ratings yet
- I Have A Headache: Jazz ChantDocument4 pagesI Have A Headache: Jazz ChantLutchie Anadia BrionesNo ratings yet
- Pasteurella MultocidaDocument9 pagesPasteurella MultocidaFatima AbasovaNo ratings yet
- Mouth Care To Reduce Ventilator Associated.22Document7 pagesMouth Care To Reduce Ventilator Associated.22ANOOPVANo ratings yet
- Ecmocard StudiesDocument34 pagesEcmocard StudiesMonica KuhonNo ratings yet
- A Study On Impact of Covid 19 OnDocument108 pagesA Study On Impact of Covid 19 OnFaisal ArifNo ratings yet
- Community Health Nursing Sample TestDocument45 pagesCommunity Health Nursing Sample TestPaulo Tiangson MejiaNo ratings yet
- 481-Article Text-2650-1-10-20231016Document6 pages481-Article Text-2650-1-10-20231016indra yanaNo ratings yet
- TOPIC 8 - Behavioral Control and Genetic Manipulation of PestsDocument11 pagesTOPIC 8 - Behavioral Control and Genetic Manipulation of PestsOliverNo ratings yet
- 2 Infection Prevention and ControlDocument5 pages2 Infection Prevention and ControlLiane BartolomeNo ratings yet
- Strawberry Leaf ScorchDocument5 pagesStrawberry Leaf ScorchJanath AnthonyNo ratings yet
- Urine Culture: D.M.M. LabDocument15 pagesUrine Culture: D.M.M. LabTomi RahmadaniNo ratings yet
- Appl 2015 Chapter 8 Health Care FacilitiesDocument16 pagesAppl 2015 Chapter 8 Health Care FacilitiesHoshyar Qadir RasulNo ratings yet