You might also like
- Understanding Delirium: Causes, Symptoms, and TreatmentDocument26 pagesUnderstanding Delirium: Causes, Symptoms, and TreatmentAnkush Sharma100% (1)
- Neurology Multiple Choice Questions With Explanations: Volume IIIFrom EverandNeurology Multiple Choice Questions With Explanations: Volume IIIRating: 4.5 out of 5 stars4.5/5 (6)
- Psychiatric Case Write Up on Brief Psychotic DisorderDocument10 pagesPsychiatric Case Write Up on Brief Psychotic DisorderBen Man JunNo ratings yet
- Delirium: DefenitionDocument18 pagesDelirium: DefenitionNanda MaulanaNo ratings yet
- Cerebral Palsy Definition, Causes, Classification and ManagementDocument25 pagesCerebral Palsy Definition, Causes, Classification and ManagementEmmanuel MukukaNo ratings yet
- Alzheimer FinalDocument121 pagesAlzheimer FinalJoanne Foja100% (1)
- Cerebral Palsy PDFDocument6 pagesCerebral Palsy PDFprasadNo ratings yet
- Understanding Dementia: Causes, Symptoms, Diagnosis and TreatmentDocument20 pagesUnderstanding Dementia: Causes, Symptoms, Diagnosis and TreatmentTurky DallolNo ratings yet
- Cerebral PalsyDocument25 pagesCerebral PalsyTINTU JOSEPHNo ratings yet
- Focal Epilepsy Causes and Clinical Features - UpToDateDocument44 pagesFocal Epilepsy Causes and Clinical Features - UpToDateBenjamínGalvanNo ratings yet
- VEM5384 Clincial Neurology DisordersDocument62 pagesVEM5384 Clincial Neurology DisordersdeadnarwhalNo ratings yet
- Classification: Epilepsy (Sometimes Referred To As A Seizure Disorder) Is A Common ChronicDocument11 pagesClassification: Epilepsy (Sometimes Referred To As A Seizure Disorder) Is A Common ChronicChristian BartolomeNo ratings yet
- Understanding Epileptic SeizuresDocument19 pagesUnderstanding Epileptic SeizuresRiki AntoNo ratings yet
- Neonatal Seizure OriginalDocument47 pagesNeonatal Seizure Originalsreekala100% (1)
- Rheumatic and Systemic Disease Clinical FindingsDocument37 pagesRheumatic and Systemic Disease Clinical FindingssallyNo ratings yet
- Factores Escoalres y EpilepsiaDocument7 pagesFactores Escoalres y EpilepsiaGabinete Psicología ÍtacaNo ratings yet
- Infantile, Childhood, and Adolescent Epilepsies.9Document34 pagesInfantile, Childhood, and Adolescent Epilepsies.9Silviana IlieNo ratings yet
- Herbal Cancer Treatment - Sabah Snake GrassDocument6 pagesHerbal Cancer Treatment - Sabah Snake GrassGelo Joson100% (2)
- Ruiz, P. - SGD and NCP On EpilepsyDocument14 pagesRuiz, P. - SGD and NCP On EpilepsyPatricia Dianne RuizNo ratings yet
- Dementia: the failing brain and its major typesDocument4 pagesDementia: the failing brain and its major typesAleja ToPaNo ratings yet
- Abdulsalam Mohammed Yakasai Physiotherapy Intervention in Children With EpiDocument17 pagesAbdulsalam Mohammed Yakasai Physiotherapy Intervention in Children With Epinaveen chaudharyNo ratings yet
- The Clinical Syndrome Cerebral MalariaDocument14 pagesThe Clinical Syndrome Cerebral Malaria1130120017 ALI ANTONO TANNo ratings yet
- Pathophysiology of Nervous System DisordersDocument9 pagesPathophysiology of Nervous System DisordersChelito GuarinNo ratings yet
- TachdjianDocument25 pagesTachdjianduracell54No ratings yet
- Down SyndromeDocument12 pagesDown SyndromeaksinuNo ratings yet
- Reversible Dementia: Sudhir SharmaDocument35 pagesReversible Dementia: Sudhir SharmaArvin PrasmanaNo ratings yet
- Sensory Impairment in Children With Medically Compromised ConditionDocument7 pagesSensory Impairment in Children With Medically Compromised ConditionAJPEDO LIFENo ratings yet
- Chapter 426 - Seizures and Epilepsy: Susan S. SpencerDocument35 pagesChapter 426 - Seizures and Epilepsy: Susan S. Spencerenriquelima19No ratings yet
- Comele EngDocument39 pagesComele EngcorneliaNo ratings yet
- 111Document12 pages111clalumarNo ratings yet
- Epilepsy in AutismDocument7 pagesEpilepsy in AutismaureliasartikaNo ratings yet
- Neurologic Diseases of RuminantsDocument6 pagesNeurologic Diseases of Ruminantsh8tcsfmmndNo ratings yet
- Dementia: Reasons of Disturbances of Higher Cerebral Functions and Their TreatmentDocument5 pagesDementia: Reasons of Disturbances of Higher Cerebral Functions and Their Treatmentjalan_zNo ratings yet
- Epilepsy and Loss of ConsciousnessDocument11 pagesEpilepsy and Loss of ConsciousnessMarwan M.No ratings yet
- Cerebral PalsyDocument8 pagesCerebral PalsyItsAHZ TimeNo ratings yet
- Effects of Cerebral Dysfunction On Neurolinguistic Performance in ChildrenDocument16 pagesEffects of Cerebral Dysfunction On Neurolinguistic Performance in Childrenhud6427No ratings yet
- Understanding Delirium: Epidemiology, Pathogenesis, Features, and DiagnosisDocument16 pagesUnderstanding Delirium: Epidemiology, Pathogenesis, Features, and DiagnosisKiara VásquezNo ratings yet
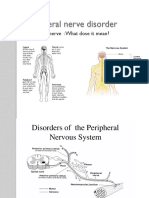
- Peripheral Nerve DisorderDocument23 pagesPeripheral Nerve DisorderSaman SarKoNo ratings yet
- Alzheimer's Disease: PathophysiologyDocument3 pagesAlzheimer's Disease: PathophysiologyMello D. ConsulNo ratings yet
- Pathology of SeizuresDocument13 pagesPathology of SeizuresZachary CohenNo ratings yet
- Erebral Alsy: A R D PDocument9 pagesErebral Alsy: A R D PMohan RajNo ratings yet
- 10 1097@NRL 0b013e31817533a3 PDFDocument10 pages10 1097@NRL 0b013e31817533a3 PDFV RakeshreddyNo ratings yet
- Cognitive and Neurologic FunctionDocument38 pagesCognitive and Neurologic Functioncoosa liquorsNo ratings yet
- 2-WS Geriatri-Dr Rensa2Document44 pages2-WS Geriatri-Dr Rensa2Yuni RochyaningsihNo ratings yet
- J Child Neurol 2001 Fennell 58 63Document6 pagesJ Child Neurol 2001 Fennell 58 63Elena GatoslocosNo ratings yet
- EpilepsyDocument2 pagesEpilepsyVenecia CalistaNo ratings yet
- Lex-Mental HealthDocument49 pagesLex-Mental HealthnasimhsNo ratings yet
- Pathophysiology 2022-2023 Exercise 14 Materials p1Document55 pagesPathophysiology 2022-2023 Exercise 14 Materials p1Luis RomanosNo ratings yet
- Pi Is 1474442205702477Document14 pagesPi Is 1474442205702477Andi Karwana CiptaNo ratings yet
- Neuromuscular System 1Document13 pagesNeuromuscular System 1Shimmering MoonNo ratings yet
- Seizures in Newborn: An Update: Review ArticleDocument6 pagesSeizures in Newborn: An Update: Review ArticleMaria Ponce de LeónNo ratings yet
- Biology: Investigatory ProjectDocument22 pagesBiology: Investigatory ProjectNarayana BaipadithayaNo ratings yet
- Diagnosis of Delirium and Confusional States - UpToDateDocument29 pagesDiagnosis of Delirium and Confusional States - UpToDateLilasNo ratings yet
- Coma and Lesser Degrees of Impaired Consciousness AreDocument45 pagesComa and Lesser Degrees of Impaired Consciousness AreAnty Dewi Sarty RNo ratings yet
- Viral Encephalitisvs ADEMDocument6 pagesViral Encephalitisvs ADEMAkdicmsNo ratings yet
- Epilepsy PDFDocument15 pagesEpilepsy PDFMohamedErrmaliNo ratings yet
- II-E Altered PerceptionDocument16 pagesII-E Altered PerceptionDharylle CariñoNo ratings yet
- Convulsion Febril PediatricsDocument11 pagesConvulsion Febril PediatricsOrlando HernandezNo ratings yet
- Introduction Ped EpilepsyDocument15 pagesIntroduction Ped EpilepsyD PharmaNo ratings yet
- EpilepsiaDocument22 pagesEpilepsiaWandaNo ratings yet
- Seizures, EpilepsyDocument8 pagesSeizures, EpilepsypertinenteNo ratings yet
- Cerebral Palsy ReviewDocument11 pagesCerebral Palsy ReviewYERNYNo ratings yet
- SUPPORTINGDocument13 pagesSUPPORTINGambiit25No ratings yet
- Childhood Seized: Linguistic Considerations of the Effects of Epilepsy on Cognition in Children with Benign Childhood EpilepsyFrom EverandChildhood Seized: Linguistic Considerations of the Effects of Epilepsy on Cognition in Children with Benign Childhood EpilepsyNo ratings yet
- Cornish Immunization Waiver PolicyDocument2 pagesCornish Immunization Waiver PolicyDonnaNo ratings yet
- 离子电渗皮肤给药系统Document9 pages离子电渗皮肤给药系统wannitan5No ratings yet
- Disease EssayDocument4 pagesDisease Essayapi-280310261No ratings yet
- Placenta Previa GuideDocument7 pagesPlacenta Previa GuideMargaret AssilasNo ratings yet
- Hiv and Aids in Children3Document25 pagesHiv and Aids in Children3Kazau FwalangaNo ratings yet
- Radiotherapy and Oncology: Karen Wong, Geoff P. Delaney, Michael B. BartonDocument5 pagesRadiotherapy and Oncology: Karen Wong, Geoff P. Delaney, Michael B. BartonFrançois IdjiwoleNo ratings yet
- Fenofibratesimvastatin Fixed-Dose Combination in TDocument13 pagesFenofibratesimvastatin Fixed-Dose Combination in TMade DeanaNo ratings yet
- Heart SurgeonDocument8 pagesHeart SurgeonayaanmalikyaseenNo ratings yet
- Community Medicine MnemonicsDocument27 pagesCommunity Medicine MnemonicsIzaz KhanNo ratings yet
- MCQ On Bleeding in Early Pregnancy: DR Manal Behery Zagazig University 2013Document36 pagesMCQ On Bleeding in Early Pregnancy: DR Manal Behery Zagazig University 2013joseph0% (1)
- Teenage Pregnancy Brief DiscussionDocument3 pagesTeenage Pregnancy Brief DiscussionJoanna Peroz100% (1)
- Seminar Electrocardiogram (ECG)Document32 pagesSeminar Electrocardiogram (ECG)Jamuna PatelNo ratings yet
- Efficacy of Varmam Therapy for Varicose VeinsDocument8 pagesEfficacy of Varmam Therapy for Varicose VeinsMurali SmatNo ratings yet
- Care of Clients With Physiologic and Psychosocial Alterations - Doc Version 1Document46 pagesCare of Clients With Physiologic and Psychosocial Alterations - Doc Version 1CLAIRE VillaviejaNo ratings yet
- Firs World Report PDFDocument35 pagesFirs World Report PDFMarv MarvNo ratings yet
- PG List 2016 17Document4 pagesPG List 2016 17Sundeep ArcheliesNo ratings yet
- Ms - CardiovascularDocument70 pagesMs - CardiovascularMark OngNo ratings yet
- Pediatrics Lecture on Malnutrition Criteria and InterventionsDocument8 pagesPediatrics Lecture on Malnutrition Criteria and InterventionskrishNo ratings yet
- Kimia Farma Per 30 September ZRMM076Document271 pagesKimia Farma Per 30 September ZRMM076SriyatiNo ratings yet
- Rekap Sisipan FormulariumDocument42 pagesRekap Sisipan FormulariumMas EmNo ratings yet
- Lecture 2 - Urine ExaminationDocument62 pagesLecture 2 - Urine ExaminationmihalachemirelaNo ratings yet
- Report WBC CountingDocument27 pagesReport WBC CountingAysha AishaNo ratings yet
- H. PyloriDocument5 pagesH. PyloriWaleed El SaidNo ratings yet
- Bril (2002) - Validation of The Toronto Clinical Scoring System For Diabetic PolyneuropathyDocument5 pagesBril (2002) - Validation of The Toronto Clinical Scoring System For Diabetic PolyneuropathyRicky WatariNo ratings yet
- Different Types of Medicines ExplainedDocument3 pagesDifferent Types of Medicines ExplainedAngelique BernardinoNo ratings yet
- Infectious Bronchitis and Its Effect On Egg Production and Egg QualityDocument2 pagesInfectious Bronchitis and Its Effect On Egg Production and Egg QualityPrima Puji RaharjoNo ratings yet