You might also like
- Carbohydrate Metabolism Mcqs for 2nd Year Mbbs Www Edu Apnafort Com TRẮC NGHIỆM HAYDocument69 pagesCarbohydrate Metabolism Mcqs for 2nd Year Mbbs Www Edu Apnafort Com TRẮC NGHIỆM HAYTrần Tiến Đạt0% (1)
- A2 Biology Notes 2016Document252 pagesA2 Biology Notes 2016MohammedKamelNo ratings yet
- Ghasem Hosseini Salekdeh (Eds.) - Agricultural Proteomics Volume 1 - Crops, Horticulture, Farm Animals, Food, Insect and Microorganisms (2016, Springer International Publishing) PDFDocument246 pagesGhasem Hosseini Salekdeh (Eds.) - Agricultural Proteomics Volume 1 - Crops, Horticulture, Farm Animals, Food, Insect and Microorganisms (2016, Springer International Publishing) PDFLa Ciencia de AlejoNo ratings yet
- Unit 5 Biology - Topic 7Document15 pagesUnit 5 Biology - Topic 7ladzville1014860100% (1)
- Lipid Movement BiochemDocument9 pagesLipid Movement BiochemCrowNo ratings yet
- NGFD 678 UjtyDocument291 pagesNGFD 678 UjtyAlfredo MunguíaNo ratings yet
- Practical 5: ENZYMES AND DIGESTIONDocument12 pagesPractical 5: ENZYMES AND DIGESTIONhafizah_9071% (7)
- Chapter8 Kinesin-13MicrotubuleDepolymerDocument17 pagesChapter8 Kinesin-13MicrotubuleDepolymer蕭雅嘉No ratings yet
- Mode of Action: InsulinDocument8 pagesMode of Action: Insulinmanus7777100% (1)
- Topic 3Document12 pagesTopic 3Ria AccadNo ratings yet
- DJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFDocument43 pagesDJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFNick_989893No ratings yet
- Cell Membrane Cell Biology and Its Application BI-1202: 2009, RRE/AB, SITH ITBDocument20 pagesCell Membrane Cell Biology and Its Application BI-1202: 2009, RRE/AB, SITH ITBFachrudin HanafiNo ratings yet
- Fgajhbsd KLMDocument17 pagesFgajhbsd KLMWira ZulfikarNo ratings yet
- Biological - MacromoleculesDocument58 pagesBiological - Macromoleculesaaronnnjohnnnn20No ratings yet
- Discussion 1 MMDocument4 pagesDiscussion 1 MMAbhishek LeveNo ratings yet
- Phosphorylation of Enoyl-Acyl Carrier Protein Reductase Inha Impacts Mycobacterial Growth and SurvivalDocument12 pagesPhosphorylation of Enoyl-Acyl Carrier Protein Reductase Inha Impacts Mycobacterial Growth and SurvivalNini FarmNo ratings yet
- Bba - Molecular Basis of Disease: SciencedirectDocument12 pagesBba - Molecular Basis of Disease: SciencedirectBiancaNo ratings yet
- He Cell 1Document34 pagesHe Cell 1Le Phuong LyNo ratings yet
- Enzyme-Associated Receptors (Insulin Receptor)Document5 pagesEnzyme-Associated Receptors (Insulin Receptor)Athena NocetoNo ratings yet
- Vitamin K: The Effect On Health Beyond Coagulation An OverviewDocument6 pagesVitamin K: The Effect On Health Beyond Coagulation An OverviewKhaerul FadlyNo ratings yet
- CHAPTER 17 The Metabolism An OverviewDocument10 pagesCHAPTER 17 The Metabolism An Overview楊畯凱No ratings yet
- Insulin Signaling PathwayshandoutDocument8 pagesInsulin Signaling PathwayshandoutManila MedNo ratings yet
- Second Messenger System in Cell SignallingDocument14 pagesSecond Messenger System in Cell SignallingMamata MishraNo ratings yet
- Bioilogy 5 - Marking SchemeDocument21 pagesBioilogy 5 - Marking Schemeclaytondc64No ratings yet
- TMP CA0 DDocument6 pagesTMP CA0 DFrontiersNo ratings yet
- It Removes A Phosphate Directly From A Substrate and Transfers It To ADPDocument8 pagesIt Removes A Phosphate Directly From A Substrate and Transfers It To ADPseussyNo ratings yet
- MIC 304-322 Metabolism 2021 - 22 Noteset 3Document16 pagesMIC 304-322 Metabolism 2021 - 22 Noteset 3kilanko timilehinNo ratings yet
- RBC MembraneDocument41 pagesRBC MembranemwambungupNo ratings yet
- PIIS0022227520392397Document16 pagesPIIS0022227520392397learningp18No ratings yet
- Cytoskeleton Molecular Motors: Structures and Their Functions in NeuronDocument10 pagesCytoskeleton Molecular Motors: Structures and Their Functions in NeuronnadaNo ratings yet
- The Kinesin KIF21B Regulates Microtubule Dynamics and Is Essential For Neuronal Morphology, Synapse Function, and Learning and MemoryDocument11 pagesThe Kinesin KIF21B Regulates Microtubule Dynamics and Is Essential For Neuronal Morphology, Synapse Function, and Learning and MemoryPeter TuNo ratings yet
- L8 2023 Protein Structure 1Document22 pagesL8 2023 Protein Structure 1maddie.jngNo ratings yet
- Biological Membranes & TransportDocument58 pagesBiological Membranes & TransportShimmering MoonNo ratings yet
- 1 (Sulfonyl) 5 (Arylsulfonyl) Indoline As PKM2 ActivatorsDocument9 pages1 (Sulfonyl) 5 (Arylsulfonyl) Indoline As PKM2 ActivatorsKa Ying Lee KatrinaNo ratings yet
- Cell MetabolismDocument10 pagesCell Metabolismbenishgulzar50No ratings yet
- Artigo 5Document4 pagesArtigo 5Uiara MariaNo ratings yet
- Energy For Muscle ContractionDocument32 pagesEnergy For Muscle ContractionAndy SkatePunkNo ratings yet
- Brain Sciences: Cofilin and Neurodegeneration: New Functions For An Old But Gold ProteinDocument12 pagesBrain Sciences: Cofilin and Neurodegeneration: New Functions For An Old But Gold Proteinjmarin55No ratings yet
- TMP A8 CCDocument13 pagesTMP A8 CCFrontiersNo ratings yet
- 2819 FullDocument9 pages2819 FullMarda AhsanyNo ratings yet
- Cell Structure and Function: Structure of The Nuclear EnvelopeDocument40 pagesCell Structure and Function: Structure of The Nuclear EnvelopeKitkat AlorroNo ratings yet
- MAP-KINASE Pathway: Intrinsic Tyrosine Kinase Activity INSULIN and Growth FactorsDocument65 pagesMAP-KINASE Pathway: Intrinsic Tyrosine Kinase Activity INSULIN and Growth FactorsUsmanNo ratings yet
- BiochemDocument5 pagesBiochemUzair MughalNo ratings yet
- Lesson 7 FA-HZDocument28 pagesLesson 7 FA-HZliaorNo ratings yet
- Post Translational Modifications On B Myosin Heavy Chain 1653708664Document24 pagesPost Translational Modifications On B Myosin Heavy Chain 1653708664Happy InsightsNo ratings yet
- Module 5. Proteins Course Outcomes: at The End of The Course, The Student Shall Be Able ToDocument6 pagesModule 5. Proteins Course Outcomes: at The End of The Course, The Student Shall Be Able ToAldine MabulacNo ratings yet
- J. Biol. Chem.-2007-Mellgren-35868-77Document10 pagesJ. Biol. Chem.-2007-Mellgren-35868-77Lie-lim CaemNo ratings yet
- Introductory One Written Answers: 1) Mention 4 Sultar Containing GAGS and The Function of Two of ThemDocument8 pagesIntroductory One Written Answers: 1) Mention 4 Sultar Containing GAGS and The Function of Two of ThemEbrahim RagabNo ratings yet
- CurrPharmDesign2007 DXR InhibitorsDocument18 pagesCurrPharmDesign2007 DXR InhibitorsVincent GeruszNo ratings yet
- Muhammad Ali ArticlesDocument87 pagesMuhammad Ali ArticlesMuhammad AliAliNo ratings yet
- Cell Cycle Checkpoints As Therapeutic Targets: Zoe A. Stewart and Jennifer A. PietenpolDocument12 pagesCell Cycle Checkpoints As Therapeutic Targets: Zoe A. Stewart and Jennifer A. PietenpolDewi Sekar MiasihNo ratings yet
- Proteins and Peptides 20msc01029Document10 pagesProteins and Peptides 20msc01029Ritu ParmarNo ratings yet
- MBB231 Chapter 9 Cytoskeleton Introduction 1197Document28 pagesMBB231 Chapter 9 Cytoskeleton Introduction 1197nathanNo ratings yet
- Protein and EnzymeDocument24 pagesProtein and EnzymeShailendraNo ratings yet
- Lipid Traffic Floppy Drives and A SuperhighwayDocument12 pagesLipid Traffic Floppy Drives and A SuperhighwayJohan AlarcónNo ratings yet
- Relating Structure To Mechanism in Creatine KinaseDocument21 pagesRelating Structure To Mechanism in Creatine KinaseAnusorn S NiiNo ratings yet
- Structure and Function of Macromolecules: 2.2.1 Carbohydrates-The Energy GiversDocument12 pagesStructure and Function of Macromolecules: 2.2.1 Carbohydrates-The Energy GiversSteven BanksNo ratings yet
- Lecture 15. Biochemistry of Muscle, Connective and Nervous Tissues, Markers of Damage, PathologyDocument46 pagesLecture 15. Biochemistry of Muscle, Connective and Nervous Tissues, Markers of Damage, PathologyВіталій Михайлович НечипорукNo ratings yet
- A Crystallographic Structure-Function Analysis of SH2 Domain Containing Inositol Polyphosphate 5-Phosphatase 2 (SHIP2)Document6 pagesA Crystallographic Structure-Function Analysis of SH2 Domain Containing Inositol Polyphosphate 5-Phosphatase 2 (SHIP2)Hani AlexNo ratings yet
- Jantos 2019Document6 pagesJantos 2019jainigNo ratings yet
- Biochemistry Anabolism of Lipids: Principal Investigator Dr. Sunil Kumar Khare, ProfessorDocument10 pagesBiochemistry Anabolism of Lipids: Principal Investigator Dr. Sunil Kumar Khare, Professorninafatima allamNo ratings yet
- Primary, Secondary, Tertiary and Quaternary Structures of A ProteinDocument21 pagesPrimary, Secondary, Tertiary and Quaternary Structures of A ProteinDALITSO CHIKOYA100% (1)
- 5 NewDocument19 pages5 Newசுப்ரமணியன் நாராயணன்No ratings yet
- Crystal Structures of Three Mercury (Ii) Complexes (HGCL 2 L) Where L Is A Bidentate Chiral Imine LigandDocument32 pagesCrystal Structures of Three Mercury (Ii) Complexes (HGCL 2 L) Where L Is A Bidentate Chiral Imine Ligandசுப்ரமணியன் நாராயணன்No ratings yet
- 21 Mehnert2002Document2 pages21 Mehnert2002சுப்ரமணியன் நாராயணன்No ratings yet
- 4 Shinjikudo2013Document17 pages4 Shinjikudo2013சுப்ரமணியன் நாராயணன்No ratings yet
- 1 Huang2019Document28 pages1 Huang2019சுப்ரமணியன் நாராயணன்No ratings yet
- 1Document16 pages1சுப்ரமணியன் நாராயணன்No ratings yet
- Evaluation of Wetting Agents by The Skein Test: Standard Test Method ForDocument3 pagesEvaluation of Wetting Agents by The Skein Test: Standard Test Method Forசுப்ரமணியன் நாராயணன்No ratings yet
- Patient Name Mr. Umesh Jain Uhid Lab No Sample Date Receiving Date 03/04/2021 11:40AMDocument1 pagePatient Name Mr. Umesh Jain Uhid Lab No Sample Date Receiving Date 03/04/2021 11:40AMAkshay ChaudhryNo ratings yet
- MICR3003 Molecular Microbiology: Molecular Basis For Expression of Multigene Phenotypes in E.coliDocument22 pagesMICR3003 Molecular Microbiology: Molecular Basis For Expression of Multigene Phenotypes in E.coliamyyma13No ratings yet
- Overview of Hemostasis - UpToDateDocument50 pagesOverview of Hemostasis - UpToDateTrung LêNo ratings yet
- Microbiology PDFDocument26 pagesMicrobiology PDFAladin herzALLAHNo ratings yet
- ADBT 3 Marker Assisted BreedingDocument48 pagesADBT 3 Marker Assisted BreedingMaham AdnanNo ratings yet
- Cell Division Cell Division: (/cellbiology/index - php/File:Historic - 1882 - Mitosis - Drawing - JPG)Document16 pagesCell Division Cell Division: (/cellbiology/index - php/File:Historic - 1882 - Mitosis - Drawing - JPG)Pinki KumariNo ratings yet
- Tet PGTRB Zoology Model Question Paper 2Document14 pagesTet PGTRB Zoology Model Question Paper 2Subbarayudu mamillaNo ratings yet
- Basic Principles of HistologyDocument10 pagesBasic Principles of HistologyDonto KonoNo ratings yet
- TT M.Sc. I SEM (C.B.C.S.) 21012020Document2 pagesTT M.Sc. I SEM (C.B.C.S.) 21012020Akash RautNo ratings yet
- CH1131 Biomolecular Engineering Week 1 Introduction 140815Document36 pagesCH1131 Biomolecular Engineering Week 1 Introduction 140815joshuaNo ratings yet
- What Is An OligoDocument4 pagesWhat Is An OligokavyamotianisgNo ratings yet
- Research Essay Final - Aidan HargettDocument10 pagesResearch Essay Final - Aidan Hargettapi-559292279No ratings yet
- Ketogenic Diet and Adipose Tissue Inflammation-A Simple Story? Fat Chance!Document2 pagesKetogenic Diet and Adipose Tissue Inflammation-A Simple Story? Fat Chance!Gabriel KeatingNo ratings yet
- Delost Introduction To Diagnostic Microbiology 390 392Document3 pagesDelost Introduction To Diagnostic Microbiology 390 392Keanne GanironNo ratings yet
- Syllabus For Biotechnology (BT) : Engineering MathematicsDocument2 pagesSyllabus For Biotechnology (BT) : Engineering MathematicsManoj SkNo ratings yet
- Chapter 1: Bacterialcell Structure, Physiology, Metabolism, and GeneticsDocument19 pagesChapter 1: Bacterialcell Structure, Physiology, Metabolism, and GeneticsNico Loko100% (1)
- Ginsenosides RC As A Novel SIRT6 Activator Protects M - 2020 - Journal of GinsDocument9 pagesGinsenosides RC As A Novel SIRT6 Activator Protects M - 2020 - Journal of GinsRafaela MirandaNo ratings yet
- Poster Jeremy WilkinsonDocument1 pagePoster Jeremy Wilkinsonsagun maharjanNo ratings yet
- Biosynthesis of Cortisol Ics-2Document3 pagesBiosynthesis of Cortisol Ics-2Kate Alyssa CatonNo ratings yet
- Molecular Docking Analysis of HER-2 Inhibitor From The ZINC Database As Anticancer AgentsDocument6 pagesMolecular Docking Analysis of HER-2 Inhibitor From The ZINC Database As Anticancer AgentsTho PhanNo ratings yet
- RnaiDocument13 pagesRnaiIrah Jane BadeNo ratings yet
- Western Blot Membrane Stripping For Restaining ProtocolDocument2 pagesWestern Blot Membrane Stripping For Restaining ProtocolDouglas SantosNo ratings yet
- Activity 10Document2 pagesActivity 10Shella Mie JamisonNo ratings yet
- Pex 03 09Document5 pagesPex 03 09Jila HafiziNo ratings yet
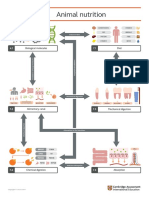
- IGCSE Biology 0610Document1 pageIGCSE Biology 0610Masroor AhmedNo ratings yet
- Life Sciences Fundamentals and Practice, Vol 1, Seventh EditionDocument16 pagesLife Sciences Fundamentals and Practice, Vol 1, Seventh EditionShalini SinhaNo ratings yet