You might also like
- Fraser’s The Behaviour and Welfare of the HorseFrom EverandFraser’s The Behaviour and Welfare of the HorseChristopher B RileyNo ratings yet
- Anthropometric, Gait and Strength Characteristics of Kenyan Distance RunnersDocument6 pagesAnthropometric, Gait and Strength Characteristics of Kenyan Distance Runnersapi-348956133No ratings yet
- Larsen Et Al-2015-Scandinavian Journal of Medicine & Science in SportsDocument9 pagesLarsen Et Al-2015-Scandinavian Journal of Medicine & Science in SportsKarliane MassariNo ratings yet
- Scandinavian Med Sci Sports - 2015 - Larsen - The Kenyan RunnersDocument9 pagesScandinavian Med Sci Sports - 2015 - Larsen - The Kenyan Runnersimmanuel1111No ratings yet
- Physiological and Performance Characteristics of Road, Mountain Bike and BMX CyclistsDocument8 pagesPhysiological and Performance Characteristics of Road, Mountain Bike and BMX CyclistsFazi Abd MalekNo ratings yet
- The Metabolic Demands of Kayaking: A Review: Jacob S. Michael, Kieron B. Rooney and Richard SmithDocument7 pagesThe Metabolic Demands of Kayaking: A Review: Jacob S. Michael, Kieron B. Rooney and Richard SmithAlexandra PredaNo ratings yet
- Most Marathon Runners at The 2017 IAAF World Championships Were Rearfoot Strikers, and Most Did Not Change Footstrike PatternDocument7 pagesMost Marathon Runners at The 2017 IAAF World Championships Were Rearfoot Strikers, and Most Did Not Change Footstrike PatternArmando NetoNo ratings yet
- Deliberate Practice in Training Differentiates The Best Kenyan and Spanish Long Distance RunnersDocument10 pagesDeliberate Practice in Training Differentiates The Best Kenyan and Spanish Long Distance RunnersJan KosteckýNo ratings yet
- 2d Endurance TrainDocument26 pages2d Endurance TrainJasunNo ratings yet
- Santos Concejero 2015Document7 pagesSantos Concejero 2015kan6de6jayNo ratings yet
- Inghametal 2008Document7 pagesInghametal 2008ko piNo ratings yet
- Locatelli The Importance of Anaerobic Glycolysis and Stiffness PDFDocument7 pagesLocatelli The Importance of Anaerobic Glycolysis and Stiffness PDFAltec AlsingNo ratings yet
- Comparing VO2max Improvement in Five Training MethodsDocument27 pagesComparing VO2max Improvement in Five Training MethodsGabriel ZahariaNo ratings yet
- Are Gait Characteristics and Ground Reaction Forces Related To Energy Cost of Running in Elite Kenyan RunnersDocument9 pagesAre Gait Characteristics and Ground Reaction Forces Related To Energy Cost of Running in Elite Kenyan RunnersMatheus Gregório KaminskiNo ratings yet
- 820 - Svedenhag 1991Document11 pages820 - Svedenhag 1991GerasimarasNo ratings yet
- Physiological Responses During Simulated 16 KM Recumbent Handcycling Time Trial and Determinants of Performance in Trained HandcyclistsDocument8 pagesPhysiological Responses During Simulated 16 KM Recumbent Handcycling Time Trial and Determinants of Performance in Trained HandcyclistsMireya CastellanosNo ratings yet
- 01 The Physiology of The World Record Holder - DR Andrew JonesDocument16 pages01 The Physiology of The World Record Holder - DR Andrew JonesbiratãNo ratings yet
- Homens e Mulheres Na Meia DistânciaDocument6 pagesHomens e Mulheres Na Meia DistânciaJúnior Alvacir CamargoNo ratings yet
- Relationship Between 800-m Running Performance and Aerobic and Anaerobic Energy Metabolism Capacities in Well-Trained Middle-Distance RunnersDocument7 pagesRelationship Between 800-m Running Performance and Aerobic and Anaerobic Energy Metabolism Capacities in Well-Trained Middle-Distance RunnersNigussie GichamoNo ratings yet
- Scandinavian Med Sci Sports - 2023 - Hanley - Changes in Running Biomechanics During The 2017 IAAF World Championships MenDocument12 pagesScandinavian Med Sci Sports - 2023 - Hanley - Changes in Running Biomechanics During The 2017 IAAF World Championships MenDeivisonNo ratings yet
- Acute Effects of Small Changes in BicyclDocument8 pagesAcute Effects of Small Changes in BicyclcsrpirataNo ratings yet
- Training Response of Adolescent Kenyan Town and Village Boys To Endurance RunningDocument10 pagesTraining Response of Adolescent Kenyan Town and Village Boys To Endurance RunningPabloAñonNo ratings yet
- Oxygen Cost and Energy Expenditure of RunningDocument7 pagesOxygen Cost and Energy Expenditure of Runningnb22714No ratings yet
- Assessment of Moderate Altitude Training On Speed Endurance Among Amateur Distance Athletes in Plateau State, NigeriaDocument6 pagesAssessment of Moderate Altitude Training On Speed Endurance Among Amateur Distance Athletes in Plateau State, NigeriaInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- HM Art 38656-10Document7 pagesHM Art 38656-10陈韦杰No ratings yet
- Performance in Heat Marino FEDocument8 pagesPerformance in Heat Marino FEMarcelo NogueraNo ratings yet
- Beattie 2016 The Effect of Strength Training For RunnersDocument16 pagesBeattie 2016 The Effect of Strength Training For RunnersRonaldo SilvaNo ratings yet
- Effects of Specific Versus Cross-TraininDocument7 pagesEffects of Specific Versus Cross-TraininManoella SilvaNo ratings yet
- Altitude Training For The MarathonDocument4 pagesAltitude Training For The MarathonbrownNo ratings yet
- The Training Characteristics of World-Class Distance Runners: An Integration of Scientific Literature and Results-Proven PracticeDocument18 pagesThe Training Characteristics of World-Class Distance Runners: An Integration of Scientific Literature and Results-Proven PracticeSportsciencesNo ratings yet
- Medical Study 1Document8 pagesMedical Study 1wfaachannel8No ratings yet
- Prediction of Maximal Oxygen Uptake in Young Adult Women JoggersDocument8 pagesPrediction of Maximal Oxygen Uptake in Young Adult Women JoggersRafaela RodriguesNo ratings yet
- Coyle Et Al - Fisiologia Ciclismo - 2005Document6 pagesCoyle Et Al - Fisiologia Ciclismo - 2005Alen_DNo ratings yet
- Art 1Document3 pagesArt 1GutoGonçalvesNo ratings yet
- 1 Grivas TSS 2020 273 117-123Document7 pages1 Grivas TSS 2020 273 117-123Yared TegegneNo ratings yet
- Vo2 MaxDocument5 pagesVo2 Maxdick headNo ratings yet
- Demandas Fisiologicas em Corridas de MontanhaDocument7 pagesDemandas Fisiologicas em Corridas de MontanhaAlan LimaNo ratings yet
- Predictors of Performance in A 4-h Mountain-Bike RaceDocument8 pagesPredictors of Performance in A 4-h Mountain-Bike RaceDanilo Lessa BernardineliNo ratings yet
- The Training of International Level Distance RunnersDocument14 pagesThe Training of International Level Distance Runnersdaniel100% (1)
- Under The Hood: Skeletal Muscle Determinants of Endurance PerformanceDocument11 pagesUnder The Hood: Skeletal Muscle Determinants of Endurance PerformanceCamilo RinconNo ratings yet
- An In-Depth Look at Running Economy: The 3 Players of Distance RunningDocument6 pagesAn In-Depth Look at Running Economy: The 3 Players of Distance RunningBryan RigginsNo ratings yet
- Combined Strength and Endurance Training in Competitive SwimmersDocument10 pagesCombined Strength and Endurance Training in Competitive SwimmersSerban ValentinNo ratings yet
- Practical Approach Altit TrainDocument8 pagesPractical Approach Altit TrainBogdan GheorghiuNo ratings yet
- Validation of An Arm Crank Ergometer Test For Use in Sedentary AdultsDocument7 pagesValidation of An Arm Crank Ergometer Test For Use in Sedentary AdultsKaribeños San JavierNo ratings yet
- Aportación 1Document12 pagesAportación 1Arturo Diaz SuarezNo ratings yet
- Hukin 69 159 PDFDocument9 pagesHukin 69 159 PDFratkopericNo ratings yet
- Icx 069Document8 pagesIcx 069Júnior Alvacir CamargoNo ratings yet
- Age and Sex Differences in Blood Lactate Response To Sprint Running in Elite Master Athlete - 20100705Document20 pagesAge and Sex Differences in Blood Lactate Response To Sprint Running in Elite Master Athlete - 20100705fzzzinNo ratings yet
- Alvero-Cruz Et Al 2019Document7 pagesAlvero-Cruz Et Al 2019people95No ratings yet
- Anaerobic and Aerobic Performance of Elite Female and Male SnowboardersDocument8 pagesAnaerobic and Aerobic Performance of Elite Female and Male SnowboardersBakhtiar WibisonoNo ratings yet
- Training Intensity Distribuition Analysis by Race Pace Vs Physiological Approach in World-Class Middle - and Long-Distance RunnersDocument24 pagesTraining Intensity Distribuition Analysis by Race Pace Vs Physiological Approach in World-Class Middle - and Long-Distance Runnersgines.gonzalez.umhNo ratings yet
- Living High-Training Low'': Effect of Moderate-Altitude Acclimatization With Low-Altitude Training On PerformanceDocument11 pagesLiving High-Training Low'': Effect of Moderate-Altitude Acclimatization With Low-Altitude Training On PerformanceDinan MitsalinaNo ratings yet
- Gasto EnergeticoDocument6 pagesGasto EnergeticoJúnior Alvacir CamargoNo ratings yet
- African Runners Dominance in Distance RunningDocument57 pagesAfrican Runners Dominance in Distance Runninglazarus81No ratings yet
- Physiology of The Elite RowerDocument8 pagesPhysiology of The Elite RowerLuky PratamaNo ratings yet
- Pedaling Confusion: Methods That Lack Sound Basis Show How Not To Train IndoorsDocument3 pagesPedaling Confusion: Methods That Lack Sound Basis Show How Not To Train IndoorsLeonardoValenzuelaNo ratings yet
- Average VO2Máx As A Function of Running Performances On Different DistancesDocument7 pagesAverage VO2Máx As A Function of Running Performances On Different DistancesCristianLopezNo ratings yet
- Farley Et Al, Journal of Strength and Conditioning Research, 2012Document6 pagesFarley Et Al, Journal of Strength and Conditioning Research, 2012api-285637348No ratings yet
- ACTN3 R577X and ACE I/D Gene Variants Influence Performance in Elite Sprinters: A Multi-Cohort StudyDocument8 pagesACTN3 R577X and ACE I/D Gene Variants Influence Performance in Elite Sprinters: A Multi-Cohort StudyJames GleickNo ratings yet
- 20230331091457-2023-Does Lactate-Guided Threshold Interval Training Within ADocument15 pages20230331091457-2023-Does Lactate-Guided Threshold Interval Training Within ARichardNo ratings yet
- Runner Muscular Balance Core Stability and Injury Prevention For Middle and Long Distance Runners PDFDocument21 pagesRunner Muscular Balance Core Stability and Injury Prevention For Middle and Long Distance Runners PDFAmmar ShamoutNo ratings yet
- Homens e Mulheres Na Meia DistânciaDocument6 pagesHomens e Mulheres Na Meia DistânciaJúnior Alvacir CamargoNo ratings yet
- E R I L A B R E M: Ffects of EST Nterval Ength On Cute Attling OPE Xercise EtabolismDocument13 pagesE R I L A B R E M: Ffects of EST Nterval Ength On Cute Attling OPE Xercise EtabolismJúnior Alvacir CamargoNo ratings yet
- Icx 069Document8 pagesIcx 069Júnior Alvacir CamargoNo ratings yet
- Esteira 1%Document8 pagesEsteira 1%Júnior Alvacir CamargoNo ratings yet
- Study of Injury and Illness Rates in Multiday.11Document7 pagesStudy of Injury and Illness Rates in Multiday.11artzaiNo ratings yet
- Hip Abductor Weakness in Distance Runners With Iliotibial Fredericson2000 PDFDocument7 pagesHip Abductor Weakness in Distance Runners With Iliotibial Fredericson2000 PDFItai IzhakNo ratings yet
- The Effect of Short-Term Swiss Ball Training On Core Stability and Running EconomyDocument8 pagesThe Effect of Short-Term Swiss Ball Training On Core Stability and Running EconomyJúnior Alvacir CamargoNo ratings yet
- Research Bites: Do Younger and Older Adults Get Similar Results With Individualized Training?Document3 pagesResearch Bites: Do Younger and Older Adults Get Similar Results With Individualized Training?Júnior Alvacir CamargoNo ratings yet
- Corrective ExercisesDocument47 pagesCorrective ExercisesZachary Lee100% (10)
- Corrective ExercisesDocument2 pagesCorrective ExercisesJúnior Alvacir CamargoNo ratings yet
- Biomechanical Analysis of The DeadliftDocument6 pagesBiomechanical Analysis of The DeadliftJúnior Alvacir CamargoNo ratings yet
- NASM Clinical Movement Analysis (PDF, 411K)Document5 pagesNASM Clinical Movement Analysis (PDF, 411K)Medina Babic100% (4)
- Tony Raynolds The Ultimate Agility Ladder GuideDocument35 pagesTony Raynolds The Ultimate Agility Ladder GuideClifton JohnstonNo ratings yet
- Comparison of The Acute Metabolic Responses To Traditional Resistance, Body-Weight, and Battling Rope ExercisesDocument11 pagesComparison of The Acute Metabolic Responses To Traditional Resistance, Body-Weight, and Battling Rope ExercisesJúnior Alvacir CamargoNo ratings yet
- Exercícios FundamentaisDocument34 pagesExercícios FundamentaisJúnior Alvacir CamargoNo ratings yet
- ABP140 mRNA Localizes To The End of Actin Cables, Independently of 3 and 5 UTRDocument1 pageABP140 mRNA Localizes To The End of Actin Cables, Independently of 3 and 5 UTRJúnior Alvacir CamargoNo ratings yet
- Certification Handbook 201505 Web RDocument72 pagesCertification Handbook 201505 Web RJose RinconNo ratings yet
- Focht Et Al-2005-Arthritis & RheumatismDocument7 pagesFocht Et Al-2005-Arthritis & RheumatismJúnior Alvacir CamargoNo ratings yet
- 7 MinutosDocument23 pages7 MinutosJúnior Alvacir CamargoNo ratings yet
- Comparison of Muscle Activation Using Various Hand Positions During The Push-Up ExerciseDocument6 pagesComparison of Muscle Activation Using Various Hand Positions During The Push-Up ExerciseJúnior Alvacir CamargoNo ratings yet
- Pa Pitch HandoutDocument12 pagesPa Pitch HandoutJúnior Alvacir CamargoNo ratings yet
- Sports: Reliability of The Single-Visit Field Test of Critical Speed in Trained and Untrained AdolescentsDocument11 pagesSports: Reliability of The Single-Visit Field Test of Critical Speed in Trained and Untrained AdolescentsJúnior Alvacir CamargoNo ratings yet
- The Effect of Short Term Swiss Ball Training On.23Document7 pagesThe Effect of Short Term Swiss Ball Training On.23Júnior Alvacir CamargoNo ratings yet
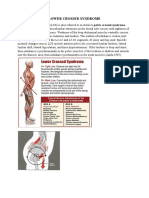
- Lower Crossed SyndromeDocument11 pagesLower Crossed SyndromeJúnior Alvacir Camargo50% (2)
- Are Deep Squats A Safe and Viable ExerciseDocument3 pagesAre Deep Squats A Safe and Viable ExerciseJúnior Alvacir CamargoNo ratings yet
- D 262Document15 pagesD 262Júnior Alvacir CamargoNo ratings yet
- 3 X 3 X 3Document21 pages3 X 3 X 3kanwalb18100% (23)
- 25 Articles To Improve Your JudoDocument79 pages25 Articles To Improve Your JudoJoão AlvesNo ratings yet
- Corrective ExercisesDocument47 pagesCorrective ExercisesZachary Lee100% (10)
- Touching The Finish Line: Predictors of Motivation Among Long Distance Elite Runners (Exploring Sports Psychology)Document12 pagesTouching The Finish Line: Predictors of Motivation Among Long Distance Elite Runners (Exploring Sports Psychology)Global Research and Development ServicesNo ratings yet
- Sprinting Ability As An Important Indicator of Performance in Elite Long-Distance RunnersDocument6 pagesSprinting Ability As An Important Indicator of Performance in Elite Long-Distance RunnersDouglas MarinNo ratings yet
- First Term Exam Phe JSS1Document2 pagesFirst Term Exam Phe JSS1Justus Ekene Nwaka100% (2)
- Marathon Training Plan - ImproverDocument6 pagesMarathon Training Plan - ImproverValmir ShalaNo ratings yet
- Explosive Power and Anaerobic Endurance For Wrestlers Andy Li-An Ho Department of Physical Education and Graduate Institute of Sports Coaching Science, Chinese Culture University, TaiwanDocument4 pagesExplosive Power and Anaerobic Endurance For Wrestlers Andy Li-An Ho Department of Physical Education and Graduate Institute of Sports Coaching Science, Chinese Culture University, TaiwanBaroes JanNo ratings yet
- Generic Information - Tata Steel Meramandali Half M - 240111 - 132218Document1 pageGeneric Information - Tata Steel Meramandali Half M - 240111 - 132218Harekrishna DasNo ratings yet
- Lesson 2 Progression and OverloadDocument2 pagesLesson 2 Progression and OverloadisaiacNo ratings yet
- ISF Official World Records Sky Running Update Oct10Document2 pagesISF Official World Records Sky Running Update Oct10Mayayo: Carrerasdemontana.comNo ratings yet
- Proposal Jakarta Marathon Pendukungan Gubernur (FA) 14082019Document25 pagesProposal Jakarta Marathon Pendukungan Gubernur (FA) 14082019lina anggraeni mariyati100% (1)
- 30 Nat PDFDocument1 page30 Nat PDFNitin SharmaNo ratings yet
- Running Events DataDocument49 pagesRunning Events DataAbhisek MohantyNo ratings yet
- Interval Training For Performance: A Scientific and Empirical PracticeDocument19 pagesInterval Training For Performance: A Scientific and Empirical Practicemattia100% (1)
- Half Marathon ProgrammeDocument5 pagesHalf Marathon ProgrammeCat HamiltonNo ratings yet
- Brainy Klasa 4 DVD Worksheet U8Document1 pageBrainy Klasa 4 DVD Worksheet U8Gabriela KarpińskaNo ratings yet
- Noakes 1990Document12 pagesNoakes 1990Einar Cumplido MartínezNo ratings yet
- Is Lactic Acid A Four Letter Word?: by Patti and Warren Finke, Team OregonDocument4 pagesIs Lactic Acid A Four Letter Word?: by Patti and Warren Finke, Team OregonАлександър Антоан НиколовNo ratings yet
- Hansons Marathon Training PlansDocument42 pagesHansons Marathon Training Plansths.it.samNo ratings yet
- Track Events and RulesDocument5 pagesTrack Events and RulesJose Karlou SantillanaNo ratings yet
- Running Training - Sprints, Middle and Long DistancesDocument3 pagesRunning Training - Sprints, Middle and Long DistancesAlejandro NavaNo ratings yet
- Atletic Finish PDFDocument11 pagesAtletic Finish PDFkareen luisa vera cabreraNo ratings yet
- Principles of Training and Conditionig NotesDocument2 pagesPrinciples of Training and Conditionig NotesBoris CastilloNo ratings yet
- What Tempo Runs and Threshold Running Are All AboutDocument1 pageWhat Tempo Runs and Threshold Running Are All AboutCharyyar BayramovNo ratings yet
- World Mountain Running Association 2018 Magazine. Official IssueDocument16 pagesWorld Mountain Running Association 2018 Magazine. Official IssueMayayo: Carrerasdemontana.comNo ratings yet
- Intermediate 2 Half Marathon PrintableDocument2 pagesIntermediate 2 Half Marathon Printableramirez.paulkennedyNo ratings yet
- IELTS Speaking Actual Tests September December 2020Document194 pagesIELTS Speaking Actual Tests September December 2020shadi shahzadiNo ratings yet
- Periodização Do Treinamento de Resistência e ForçaDocument39 pagesPeriodização Do Treinamento de Resistência e ForçaThiago Oliveira100% (1)
- BerlinDocument1 pageBerlinLive SportsNo ratings yet
- Bahir Dar University Sport Academy: PHD Program in Athletics CoachingDocument15 pagesBahir Dar University Sport Academy: PHD Program in Athletics Coachingdemeku akaluNo ratings yet
- The Training of International Level Distance RunnersDocument14 pagesThe Training of International Level Distance Runnersdaniel100% (1)
- MarathonDocument15 pagesMarathonapi-3820304No ratings yet