Professional Documents
Culture Documents
Parental Investment in Offspring in Variable Environments: Theoretical and Empirical Considerations
Uploaded by
Andre LanzerOriginal Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Parental Investment in Offspring in Variable Environments: Theoretical and Empirical Considerations
Uploaded by
Andre LanzerCopyright:
Available Formats
See discussions, stats, and author profiles for this publication at: https://www.researchgate.
net/publication/249140236
McGinley, M. A. , Temme, D. H. & Geber, M. A. Parental investment in
offspring in variable environments: theoretical and empirical considerations.
Am. Nat. 130, 370-398
Article in The American Naturalist · September 1987
DOI: 10.1086/284716
CITATIONS READS
543 1,061
3 authors, including:
Mark Mcginley Monica A Geber
Lingnan University Cornell University
16 PUBLICATIONS 1,265 CITATIONS 103 PUBLICATIONS 8,769 CITATIONS
SEE PROFILE SEE PROFILE
Some of the authors of this publication are also working on these related projects:
Adaptation, demography, and range View project
Ashmeadiella phylogeny View project
All content following this page was uploaded by Monica A Geber on 09 March 2014.
The user has requested enhancement of the downloaded file.
The University of Chicago
Parental Investment in Offspring in Variable Environments: Theoretical and Empirical
Considerations
Author(s): Mark A. McGinley, David H. Temme and Monica A. Geber
Source: The American Naturalist, Vol. 130, No. 3 (Sep., 1987), pp. 370-398
Published by: The University of Chicago Press for The American Society of Naturalists
Stable URL: http://www.jstor.org/stable/2461891 .
Accessed: 09/03/2014 18:02
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .
http://www.jstor.org/page/info/about/policies/terms.jsp
.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of
content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms
of scholarship. For more information about JSTOR, please contact support@jstor.org.
The University of Chicago Press, The American Society of Naturalists, The University of Chicago are
collaborating with JSTOR to digitize, preserve and extend access to The American Naturalist.
http://www.jstor.org
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
Vol. 130,No. 3 The American
Naturalist September
1987
PARENTAL INVESTMENT IN OFFSPRING
IN VARIABLE ENVIRONMENTS:
THEORETICAL AND EMPIRICAL CONSIDERATIONS
MARK A. MCGINLEY, DAVID H. TEMME, AND MONICA A. GEBER
Department
ofBiology,University
of Utah,SaltLake City,Utah84112
SubmittedSeptember25, 1986; Accepted February13, 1987
The studyoflifehistorycharacteristics involvesprimarily thestudyof trade-
offs.One of the mostfundamental trade-offs is thatbetweenthe amountof
resourcesinvestedperoffspring and thenumberofoffspring produced.Parental
investment mayoftenbe criticalto the survivaland growthof young,but the
largertheinvestment peroffspring, thefewerthenumber thatcan be
ofoffspring
produced.In a theoretical analysisofthistrade-off, C. Smithand Fretwell(1974)
concludedthatan optimalinvestment level existsand thatparentalfitnessis
maximizedby investing equallyin all young.
The Smith-Fretwell modelhas beencriticized becauseitdoes notconsiderthe
effectof environmental variationon offspring investment. For example,Janzen
(1977a)proposedthatbecause seeds fallintoa spatiallyheterogeneous environ-
ment,thereis no oneoptimalseedsize buta rangeofoptimalsizesthatvarieswith
thehabitat.He suggestedthatnaturalselectionfavorsan optimaldistribution of
seed sizes withina plant'sseed crop.Similarly, Capineraarguedthattheoptimal
propagulesize varies withenvironmental conditionsand concludedthatthe
Smith-Fretwell model"appears to have littlevaliditywithregardto plantsand
insects" (1979, p. 358). Recently,R. Kaplan and Cooper arguedthat "it is
becomingincreasingly clearthattheidea of optimaleggsize is hardto defend"
(1984,p. 393).The variability
inoffspring size hasbeenviewedas an adaptationto
variableenvironments in insects(Capinera1979),isopods (Brodyand Lawlor
1984),salamanders (R. Kaplan 1980),frogs(Crump1981,1984),and plants(Jan-
zen 1977b;Capinera1979;Stampand Lucas 1983;Thompson1984).
In thispaper,we examinethe effectof spatialand temporalenvironmental
variabilityon theoptimaloffspring size(s) through theuse ofmathematical mod-
els. We findthatenvironmental variability does notnecessarilyfavortheproduc-
tionof a rangeof offspring sizes by a parent.Most often,parentalfitnessis
maximizedby investing equallyin all young.Considerablevariationin offspring
size has, however,been documented forbothplantsand animals(see below);in
theDiscussion,we considerfactorsthatmaypreventan organism frommaking
equal-sizedyoung.
Am. Nat. 1987. Vol. 130, pp. 370-398.
? 1987 by The Universityof Chicago. 0003-0147/87/3003-0003$02.00.
All rightsreserved.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 371
A B
FIG.
1.,Terlatonsipetweood good
bad
good
and badenvironmentbad
Sb
m Km
size (8)
Offspring size (8)
Offspring
FIG. 1.-A, The relationshipbetween offspring fitnessand size in environmentsof good
and poor quality. The optimal offspringsizes (&g and Sb) inl the two environmentsare
determinedgraphically.B, The relationshipbetween parentalfitnessand offspringsize in
good and bad environments.
THE SMITH-FRETWELL MODEL
Much of the disagreement over the optimalparental-investment strategy ap-
pearsto stemfromconfusionabouttheSmith-Fretwell model(see theDiscus-
sion). C. Smithand Fretwell(1974)madetwo assumptions concerning therela-
tionshipbetweenthe amountof investment per offspring and offspring fitness.
First,a minimum levelofinvestment is requiredifan offspring is to surviveat all.
Second, above thisthresholdlevel, the relationship betweenthe resourcesin-
vestedin an offspring and offspringfitnessis positiveand convex(fig.IA).
Undertheseassumptions, a singleoptimallevelof investment maximizespa-
rentalfitness.
Thiscan be determined graphically (see fig.1),anditis thelevelthat
maximizesoffspring fitnessperunitofresourceinvested.Unlesstherelationship
betweenoffspring size and offspring fitnessis bimodal,equal investment in all
youngrepresents thetheoretical optimalstrategy.
The offspring-fitness curvecan easilybe converted intoa parental-fitness curve
bymultiplying fitness perinvestedresource(slopeofthetangent to theoffspring-
fitnesscurve,fig.1A)bytheamountofresourcesavailableto a parent(fig.iB). If
offspring fitnessincreaseswithoffspring size, parentsinvesting less thanthe
optimallevel have a lowerfitnessthanthoseinvesting more(fig.iB).
THE MODEL
To modelthesimplestof variableenvironments, we examinean environment
betweeninvestment
withonlytwostates:good (g) and bad (b). The relationship
peroffspring and offspringfitness betweenthetwoenvironments
differs because
of bioticor abioticfactors.We assume,in particular,thattheminimum viable
offspringsize is smallerin the good environmentthanin thebad environment
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
372 THE AMERICANNATURALIST
because an offspringshouldrequirefewerresourcesto becomeestablishedin a
morefavorablesite (fig.IA). We also assumethat,fora givensize, offspring
fitnessis greaterin the good environmentthanin the bad environment. We
describethe relationshipbetweenoffspringfitness(F) and offspring
size (s) as
follows:in thegood environment,
Fg(s) =1 -(mls)a s > m,
Fg(s) = 0 otherwise
and in thebad environment,
Fb(s) = 1 (kmls)a s > km,
Fb(s) = 0 otherwise
wherem is theminimum viableoffspring size abovewhichF > 0; k is a constant
thatspecifiesthedifference inqualitybetweenthetwoenvironments (k > 1); and
a, also a constant,
determines therateofapproachoftheoffspring-fitness curveto
itsasymptote. Withk > 1, theminimum viableoffspringsize is largerin thebad
environment (km),andtherateofapproachofF to itsasymptotic valueis slower,
comparedwiththegood environment (fig.1A).
The optimaloffspring sizes (s) in the two environments can be determined
analytically dFgids ordFbldsequaltotheslopeofthetangent
bysetting linetothe
curveand solvingfors. The optimalinvestment levelsareas follows:inthegood
environment,
s= m(l + a)l/a;
and in thebad environment,
Sb= km(l + a)l/a.
Notethatwhenk > 1,S^bis largerthans^g(see fig.1A).Throughout thispaper,we
referto theoptimaloffspringsizes in thegoodandbad environments as "small"
and "large,"respectively.
We use thisrepresentation ofenvironmental variability
to consideroptimalpatternsofinvestment inthreesituations: in
spatialvariation
theenvironment withno densitydependenceon offspring spatialvariation
fitness;
withdensity-dependentreductions in offspring
fitness;and temporalvariation.
Spatial Variability
In a spatiallyvariableenvironment, ifa parentmakesall of its offspring
the
samesize, whatis theoptimalsize, s? Andcan parentsachievehigherfitness by
producing a combination of offspring
sizes ratherthanby producing equal-sized
offspring?
We assumethatthetwohabitattypes,goodandbad, occurintheproportion p
and 1 - p, respectively, and offspringare placedrandomly acrossthehabitats.
Parentalfitness(W) is a function of the numberof offspringproducedand the
fitnessof each offspring.
If all offspring
are the same size, the numberof offspring produced(N) is
=
N(s) Els, whereE is theresourceavailableforreproduction.Parentalfitness
in
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 373
co
co 10 _
C
.0~~~~S
a.) \
sm a=1I2=
co I I
0.1 0.5 0.9
Frequency of good habitat (p)
FIG. 2.-Phase diagram depictingtheoptimaloffspring
size as a function
ofp, thefre-
quencyof good habitats,and of k, the difference
in qualitybetweenthe good and bad
habitats,forseveralvalues of a, a measureof how quicklyoffspring fitnessreachesan
asymptote as size increases.
a spatiallyvariableenvironment
can be written
as
W(s) = pN(s)Fg(s) + (1 - p)N(s)Fb(s). (1)
Note that if m < s < km, then Fb = 0 and equation (1) collapses to W(s) =
pN(s)Fg(s).
Wills^everbe less thankm;thatis, shoulda parenteverproduceyoungso small
thatit gainsno fitnessfromoffspring landingin thepoorhabitat?If theloss in
parentalfitness
fromoffspring dyinginthepoorhabitatis morethanoffset bythe
gainstoa parentfrommaking moreoffspring, a parentshouldmakeyoungsmaller
thankm.Thisoccurswhenever
k > [(1 - pl+a)Ipa(1 - p)]l/a and s^g< km (2)
(AppendixA). Furthermore, the optimaloffspring size is s^g.In otherwords,
whenever thedifference
inquality(k)betweenthetwohabitatsis largeand/orthe
environment consistsmostlyofthegoodhabitattype(p large),a parentdoes best
by producing onlysmalloffspring (fig.2). If condition(2) is reversed,a parent
shouldmakeoffspring largerthanthe minimum viablesize forthebad habitat
(s > km). The optimalsize is
Sm= m(1+ a)l/a [p + (1 - p)ka]lla
(see fig.2). Note thatS^mis intermediate
in size betweensg and Sb. Thus,in a
spatiallyheterogeneous environment,
theoptimaloffspringsize,s^,mayrepresent
a compromise betweenwhatis optimalin theseparatehabitats.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
374 THE AMERICANNATURALIST
Can a parentever do betterby producing offspringof two sizes ratherthan
making themall thesamesize? The argument hereis thata parentmight benefitif
some offspring of the "correct" (optimal)size dispersedto the appropriate
habitat.
The setofall possibletwo-sizestrategiesis clearlyenormous, but,fortunately,
we neednotconsiderall ofthem.Rather,itcan be shownthatthesetoftwo-size
strategies
maximizing parentalfitnessconsistsof combinations of small(sg) and
medium-sized (S^m)young(see AppendixB). The fitnessof a parentwho makes
somecombination ofthesetwooffspring sizes can be writtenas
W(Qg9Sm)= pqN(S^g)Fg(s^g) + p(l - m)
q)N(S^m)Fg(S
+ (1 - p)(l - q)N(S^m)Fb(S^m)+ (1 - p)qN(S^g)Fb(S^g),
whereq is thefraction of parentalresources(E) allocatedto theproductionof
smalloffspring.
We find,forall valuesof k andp, thatthefitness ofa parentusinga two-size
is alwaysintermediate
strategy betweenthefitness ofa parentmakingonlysmall
(sg) and thatof a parentproducingonly mediumoffspring
offspring (S^m).Indeed,
> W(Sfm) when
W(S^g)> W(S^gsg^m) k > [(1 - pl+a)Ipa(1 _ p)]l/a
(i.e., wheneverthegood habitatis widespreadand/oris of muchbetterquality
thanthepoorhabitat),and conversely,
W(Qg) < WQ9g,1m)< W(fim) when k < [(1 -pl+a)Ipa(l - p)]l/a
(i.e., whenever thegoodhabitatis rareand/or does notdiffer muchinqualityfrom
the poor habitat).In conclusion,underrandomdispersalof offspring, spatial
variation neverfavorsvariationin offspring size.
Whatifparentscan directtheiroffspring to theappropriate habitatmoreoften
thanwouldbe predictedby randomdispersal?If a parenthad perfectcontrol,it
shoulddirectall offspring to the good habitatsinceoffspring fitnessis always
higher there;and,furthermore, itshouldmakeonlysmall(sg) offspring. The same
wouldapplyifoffspring couldchoosetheappropriate habitat.
Ifparentshaveless thanperfect controloverdispersal,a fraction cp rather than
p of the reach
offspring the good habitat
(1 < c < l Ip). In ourmodels, cp shouldbe
substituted forp; thischangesneithertheformoftheequationsnorthegeneral
predictions of the model.Equal investment in all offspring is stillthe optimal
parentalstrategy. Nonrandom dispersaltothefavorable habitattendsto decrease
theoptimaloffspring size.
Fitness
Spatial Variationand Density-DependentOffspring
In theprecedingmodel,we assumedthatthefitnessof an offspring depends
onlyon its size and on environmentalqualitybutis independent
of thenumber
(density)ofotheryoungsharing thehabitat.We thenshowedthatparentsproduc-
ingonlysmalloffspring themto thegood habitathave thehighest
and directing
Ifall members
fitness. ofthepopulation adoptedthisstrategy,
however,offspring
densityin thegood habitatwouldbecomeveryhigh.If, as is morereasonable,
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 375
offspring fitnessis density-dependent, increasing densitiesof youngin thegood
habitatwillreduceenvironmental qualitythereuntilfitnesses areequalizedacross
bothhabitats.Naturalselectionmight thenfavorparentsthatproducedbothlarge
and smalloffspring and directedtheiroffspring to theappropriate environments.
We now considertheeffectof densitydependencein offspring fitnesson the
optimalparentalinvestment strategy. Here,thefitness associatedwitha particu-
larparentalstrategy dependson thebehaviorofotherparentsinthepopulation.
We therefore use thetoolsofgametheory tofindtheevolutionarily stablestrategy
(ESS) ofparentalinvestment. The ESS investment is thestrategy thatcannotbe
invadedby anyotherstrategy parentsmightadopt.
We considertwoparentalinvestment strategies:SI, theproduction ofa single
offspring size thatmaximizesparentalfitness underrandomdispersal(i.e., s = g
orSm);andS2,theproduction ofa combination oflargeandsmalloffspring whom
parentsattempt to directto thecorrecthabitats.
To findtheESS, we compare,through theuse of computersimulations, the
expectedfitness ofa rareindividual usingstrategy Si ina population consistingof
individualsusingstrategy Sj (i, j = 1, 2; i ?7j) and theexpectedfitnessof an
individual usingSi in a populationusingSi (i = 1, 2) (see MaynardSmith1982,
chap. 2). These fitnessrelationships are developedexplicitly in AppendixC.
In our simulations, we variedq, thefraction of resourcesallocatedto small
offspring by parentsusing strategyS2; h, the abilityof S2 parentsto direct
offspring appropriately to the two habitats;c, the intensity of the density-
dependent fitnessloss to offspring; and k, thedifference in qualitybetweenthe
twohabitats.
We foundthattheproduction ofvariableoffspring sizes (strategyS2) is favored
undera verylimitedsetofconditions (table1). Underrandomdispersal,a single
offspring size (eithersg or Sm)is alwaysoptimal.As parentalabilityto direct
offspring to the appropriate habitatimproves,therangeof conditions favoring
variableoffspring sizes increases.Nonetheless,even withperfectcontrolover
offspring dispersal,parentsare not alwaysselectedto makevariableoffspring
sizes. This occursonlywhenthe intensity of densitydependencein offspring
fitnessis greatenoughthattherareparentdirecting largeoffspring to thebad
habitatdoes betterthanparentssendingsmalloffspring to thegoodhabitat.The
levelofdensity-dependent fitness loss necessaryto favortheproduction ofvari-
able offspring sizes increasesas the disparity in the qualitybetweenthe two
habitatsbecomesgreater.
Temporal Variability
Within-year variability.-Iftheenvironment variesduring thebreeding season,
parentalfitnessis maximizedby adjustingoffspring size to theprevailing condi-
tions.Within-year environmental variability
couldtherefore lead to selectionfor
offspringsize variationifparents can assess environmental conditions and pro-
duce youngoftheappropriate size.
Between-year variability.-When environmentalconditions vary between
years,itis no longercorrectto use thearithmetic
meanoffitness as a measureof
long-term success(Cohen 1966;Lacey et al. 1983).Rather,long-termsuccessis
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
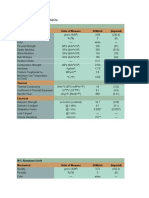
TABLE 1
OPTIMAL INVESTMENT PER OFFSPRING IN A SPATIALLY VARIABLE ENVIRONMENT WITH
DENSITY-DEPENDENT REDUCTION IN OFFSPRING FITNESS
PROPORTION OF LEVEL OF DENSITY-DEPENDENT FITNESS LOSS
RESOURCES TO
SMALL OFFSPRING .0000 .0002 .0004 .0006 .0008 .0010
o.6 DISPERSED CORRECTLY
k = 1.5
0 M M M M M M
0.2 M M M M M M
0.4 M M M M M M
0.6 M M M M M M
0.8 M M M M M M
1.0 M M M M M M
k = 2.5
0 M M M M M M
0.2 M M M M M M
0.4 M M M M M M
0.6 M M M M M M
0.8 S S SM SM SML SML
1.0 S S SM SM SML SML
k = 3.5
0 S S S S S S
0.2 S S S S S S
0.4 S S S S S S
0.6 S S S S SL SL
0.8 S S S SL SL SL
1.0 S S S SL SL SL
k = 4.5
0 S S S S S S
0.2 S S S S S S
0.4 S S S S S S
0.6 S S S S SL SL
0.8 S S S S SL SL
1.0 S S S S SL SL
o.8 DISPERSED CORRECTLY
k = 1.5
o M M M M M M
0.2 M M M M M M
0.4 M M M M M M
0.6 S S SL SL SML SML
0.8 S S SL SML SML SML
1.0 S S SML SML SML SML
k = 2.5
0 M M M M M M
0.2 M M M M M M
0.4 S S S SL SL SML
0.6 S S S SL SL SL
0.8 S S S SL SML SML
1.0 S S S SML SML SML
k = 3.5
0 S S S S S S
0.2 S S S S S S
0.4 S S S S SML SML
0.6 S S S S SL SL
0.8 S S S S SL SL
1.0 S S S S SL SL
376
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
TABLE 1 (Continued)
PROPORTION OF LEVEL OF DENSITY-DEPENDENT FITNESS Loss
RESOURCES TO
SMALL OFFSPRING .0000 .0002 .0004 .0006 .0008 .0010
o.8 DISPERSED CORRECTLY (CONTINUED)
k = 4.5
0 S S S S S S
0.2 S S S S S S
0.4 S S S S S S
0.6 S S S S SL SL
0.8 S S S S SL SL
1.0 S S S S SL SL
PERFECT DISPERSAL
k = 1.5
0 M M M M M M
0.2 M M M M M M
0.4 M M M M M SL
0.6 S S SL SL SL SL
0.8 S S SL SML SML SL
1.0 S S SML SML SML SML
k= 2.5
0 M M M M M M
0.2 M M M M M M
0.4 S S SL M M SL
0.6 S S SL SL SL SL
0.8 S S SL SML SML SML
1.0 S S SML SML SML SML
k = 3.5
0 S S S S S S
0.2 S S S S S S
0.4 S S S SL SL SML
0.6 S S S SL SL SL
0.8 S S S SL SL SL
1.0 S S S SL SL SL
k = 4.5
0 S S S S S S
0.2 S S S S S S
0.4 S S S SML S SML
0.6 S S S SL SL SL
0.8 S S S SL SL SL
1.0 S S S SL SL SL
NOTE.-The optimaloffspringsize depends on the proportionof resources allocated to large and
small offspring by parentsproducingboth offspring sizes, the level of density-dependentfitnessloss
(c), the differencein qualitybetween habitats(k), and the abilityto directoffspring
to the appropriate
habitat(h). Because the simulationsassumed that parentsrandomlydirectingtheiroffspringwould
place 50% in each habitat,parentswho were able to disperse75% of theiroffspring correctlywere 50%
betterat directingtheir offspringto the appropriatehabitat. S, small offspring;M, medium-sized
offspring;L, large offspring.
377
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
378 THE AMERICANNATURALIST
betterreflected eitherbythegeometric meanoffitness (forstrictly annualorgan-
isms) or by the arithmetic mean discountedforthe varianceof fitnessacross
years.We use bothfitness measuresto evaluatetheoptimalparentalinvestment
strategy.
Usingthegeometric meanoffitness, R. KaplanandCooper(1984)modeledthe
effectsof year-to-year environmental variationon parentalinvestment in off-
spring,and theyconcludedthattheproduction of variableoffspring is favored.
The sortof variationin offspring size thattheseauthorsconsider,however,is
variationin the sizes of offspring betweenyears(clutches)ratherthanwithin
years.Thus,theycontrasttwoparentalstrategies in theirmodel:a parentpro-
duces offspring ofequal size withina year(clutch)and thesame-sizedoffspring
everyyear;andoffspring size is constant within a year(clutch)butvariesbetween
years(clutches).We feelthatwithin-year variationin offspring size is consider-
able in bothanimalsand plants,and it is thistypeof size variationthathas
prompted argument abouttheadaptivesignificance ofoffspring size variation.
We also thinkthatKaplanandCooper'sconclusion thatvariable-sized offspring
arefavoredrestson a particular, andperhapsinappropriate, assumption aboutthe
relationships amongoffspring fitness, offspring size, and environmental quality.
In discussingtheirmodel,theyfailedto stateexplicitly how offspring fitness
dependson size and environment, butthisrelationship can be deducedfromthe
relationship amongparentalfitness, offspringsize,andenvironmental quality(see
fig.3A). Theyassumedthatforeach environment i, a singleoffspring size, si,
existsthatmaximizesparentalfitness;and thatthefitnessof parentsproducing
offspring ofsize si declinesas theenvironment eitherdeteriorates orimprovesin
qualityrelativeto environment i. Since parentalfitnessis simplytheproductof
offspring fitnesstimesthe numberof offspring produced,thepreviousassump-
tionsabout parentalfitnessmustapplyto offspring as well; namely,foreach
environment i, offspringfitnessis maximum at somesize si; as theenvironment
eitherworsensor improvesin quality,offspring ofsize si havelowerfitness (fig.
3B). It is morelikely,we think,thatbothoffspring fitnessand parentalfitness
alwaysincreaseas environments improveand decreaseas environments deterio-
rate,regardlessofoffspring size, and thatlargeroffspring do betterthansmaller
offspring in all environments (figs.3C,D). If theselatterassumptions are used in
KaplanandCooper'smodel,between-clutch (yearly)variation inoffspring size is
no longerthefavoredparentalstrategy.
Because ofthesedifficulties withKaplanandCooper'smodel,we investigated
further theeffect ofyear-to-year environmental variation on parentalinvestment.
Usingcomputer simulations,we comparedthefitness ofthreeparentalstrategies:
(1) parentsare capable of predicting environmental qualityand of adjusting
offspring size accordingly; (2) parentsproduceonlyone offspring size everyyear
(eithersg Sm, or sb); and (3) parentsmakea combination ofbothsmallandlarge
offspring each year.
In oursimulations, we variedthedifference inqualitybetweenyears(k)andthe
frequency ofgoodyears(p). We usedboththegeometric meanandthearithmetic
meandiscountedby the variance(mean - variance/[2x mean];Lacey et al.
1983)as measuresoffitness.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 379
Kaplan & Cooper 1984
A X-B
U) C~~~~~~~~~~~U
_- small med large
large
med
i) ci small
co
0~ ~~~~~~
good bad good bad
Environmental
quality Environmental
quality
This Paper
C D
0~~~~~~~~~~~~~~~~0
CD
CO) ~~~~~~~~med
co
entprdcn
0 small,mdu-ie,olagofsrn,aasuebyRKpanndCpr
sml med lrge
~~~~~~~~~~~~~~~~~large
good bad good bad
tha wudrslfrmteprna-insfnctodecidinAC,Terlinsp
Environmental quality Environmental quality
FIG.3.-A, The relationship betweenparentalfitness qualityforpar-
and environmental
entsproducing small,medium-sized, as assumedbyR. KaplanandCooper
orlargeoffspring,
(1984,fig.6, p. 408).B, The relationship
betweenoffspring fitness
andenvironmentalquality
thatwouldresultfromthe parental-fitness functiondescribedin A. C, The relationship
betweenoffspring fitnessand environmental qualityforsmall,medium-sized, and large
as assumedinthispaper.D, Therelationship
offspring, betweenparentalfitness
andenviron-
mentalqualityforparentsproducing small,medium-sized, derivedfromC.
orlargeoffspring,
Parentsadoptingstrategy 1 abovealwayshadthehighest fitness.Whenparents
wereunableto predictenvironmental theoptimalstrategy
conditions, depended
on bothk andp and on thechosenfitness measure(table2).
Whenfitness is measuredbythearithmetic meandiscounted bythevariance,a
singleoffspringsize is generally thefavoredinvestment strategy.As withspatial
smalloffspring
variability, (kg) are optimal
whengood years(sites)are common
and/orwhenthedifference betweenthequalityof goodand bad years(sites)is
large.In contrastto resultswithspatialvariability, largeoffspring(Sb) are fre-
quentlyfavoredover medium-sized (Sm) because the variancein off-
offspring
springfitnessis less forlargethanformedium-sized progeny.
Whenthe geometric meanof fitnessis used, theoptimalparentalstrategy is
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
380 THE AMERICAN NATURALIST
TABLE 2
RESULTS OF SIMULATIONS CALCULATING THE OPTIMAL OFFSPRING SIZE IN A
TEMPORALLY VARIABLE ENVIRONMENT
DIFFERENCE IN QUALITY BETWEEN YEARS (k)
2 3 4 6 8
PROBABILITY
OF A GOOD YEAR G A G A G A G A G A
0.2 M M M L M = L L M==L L M = L L
0.5 M M M M L L V V V S
0.8 M M V S V S V S V S
NOTE.-Different values of the probabilityof a good year and the differencein quality between
years are shown using two measures of fitness:the geometricmean (G) and the arithmeticmean
discounted for variance (A). S, small offspringonly; M, medium-sizedoffspringonly; L, large
offspringonly; V, both large and small offspring.
oftenthesame,exceptthatvariableoffspring sizesare nowfavoredundercertain
circumstances (table2). In particular,
whenever smalloffspringarefavoredbythe
arithmetic fitnessmeasure,variableoffspring are optimalwhenthe geometric
meanoffitness is used. Thisis becausethegeometric meanheavilyweightsyears
withlow reproductive success, as wouldoccurwhenonlysmalloffspring are
producedin a bad year.Indeed,ifreproduction failscompletelyin anyyear,the
geometric meanof fitnessequals zero. This is equivalentto theextinction of a
lineagein a strictlyannualorganism.
Year-to-year environmental variationcouldthenfavoroffspring size variation
in strictly
annualspecies.In manyannualplantsand animals,however,perenni-
alityis achievedthrough delayedgermination ordormancy (Cohen1966;Venable
and Lawlor1980).In is certainly premature,giventheresultsofthesemodels,to
arguethattemporal variabilityalwaysfavorstheproduction ofvariableoffspring.
DISCUSSION
We have foundthatthe optimalsolutionto the trade-off betweensize and
numberof offspring is mostoftenequal investment in each offspring.
Consider-
able variationin offspringsize has been found,however,in bothanimalsand
plants.Thislackoffitbetweenprediction and observation couldariseforseveral
reasons.Here, we discussthe importance of the models'assumptions forthe
predictionof equal investment. We thenconsiderthe role of developmental
constraintsin preventingorganisms frominvesting equallyin offspring
and dis-
cuss how effective canalizingselectionfora uniform offspringsize mightbe in
variableenvironments. Finally,we outlineconditionsunderwhichvariationin
offspringsize couldbe adaptive.
Importance of Assumptions
In usingtheoretical models,we are forcedto stateour assumptionsand to
formalize ourlogic.In theirmodelofparentalinvestment,
C. Smithand Fretwell
(1974) assumeda constantenvironment, convexrelationship
an increasing be-
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 381
TABLE 3
STUDIES SHOWING POSITIVE RELATIONSHIPSBETWEENSEED SIZE AND SEEDLING CHARACTERISTICS
Seedling
Characteristic Sources
Size, biomass 1945;Fowells1953;Peace 1953;
Oexemann1941;Spurr1944;Righter
Rogler 1954; Kneebone 1955; Kneebone and Cremer 1955; Black
and
1956;Langdon1958;BeveridgeandWilsie1959;Kaufmann
andWassom1963;Cope 1966;Twamley
McFadden1960;Stickler
1967; W. Williams 1967; Autensonand Walton 1970; Peacock and
Hawkins 1970; Boyd et al. 1971; Carletonand Cooper 1972; Lowe
and Ries 1973; Ries and Everson 1973; Wulff1973; Ayers et al.
1976; Evans and Bhatt 1977; Lewis and Garcia 1979; Bulisani and
Warner 1980; Jacobsohnand Globerson 1980; Howe and Richter
1982; Pitelka et al. 1983; Glewen and Vogel 1984; Gross 1984;
Morse and Schmitt1985; Marshall 1986
Leaf area, size Black 1956; Kalton and Christie1957; R. Thomas 1966; Austinand
Longden 1967; Harper and Obeid 1967; W. Williams 1967; Malik
and Kanwar 1969; Wulff1973; Jacobsohnand Globerson 1980;
Cideciyan and Malloch 1982; Howe and Richter1982
Leafnumber Black 1956
Cotyledonsize Stanton1985
Rootsize Austinand Longden1967;Malikand Kanwar1969;Lowe andRies
andGloberson1980
1973;0. Smithet al. 1973;Jacobsohn
Seed yield Kalton and Christie1957; Demirbicakmaket al. 1963; Cope 1966;
Stanton 1984a, 1985
Germination
% Spurr 1944; Kneeboneand Cremer1955; Ackermanand Gordon 1969;
Cideciyan and Malloch 1982; Nelson and Johnson1983; Pitelka et
al. 1983; Gross 1984; Hendrix 1984; Morse and Schmitt1985
Speed ofgermination Spurr1944;Malikand Kanwar1969;Weis 1982
Emergence % Rogler1954;Kneeboneand Cremer1955; Sticklerand Wasson 1963;
Dolan 1984
Speedofemergence Kneeboneand Cremer1955
Depthofemergence Black 1956
Survival Oexemann1941;Spurr1944;Morseand Schmitt
1985
tweenoffspring fitnessand size, and limitedresourcesforreproduction. Their
assumptions andconsequently theirprediction ofequalinvestment inyounghave
been criticized.
Here,we have shownthatrelaxingtheassumption ofa constantenvironment
does not necessarilylead to selectionforvariableinvestment in offspring.Re-
searcherswho have arguedthatenvironmental variationfavorsoffspring size
variationeitherhave notmadea formalanalysisoftheirargument or haverelied
on whatwe considerinappropriate assumptions aboutthe relationships of off-
springsize to offspringfitnessand environmental quality.
Capinera(1979)argued,and R. KaplanandCooper(1984)implied(see above),
thatoffspring fitnessmaynotalwaysincreasewithincreasesin parentalinvest-
mentbutmight be maximized at someintermediate size. In plants,however,there
is overwhelming evidencefora positivecorrelation betweenseed size and plant
characteristicsrelatedto fitness(table3). Furthermore, recentstudieson species
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
382 THE AMERICAN NATURALIST
TABLE 4
STUDIES SUGGESTING INCREASING CONVEX RELATIONSHIPS BETWEEN SEED SIZE AND SEEDLING
CHARACTERISTICS
Seedling
Characteristic Sources
Seedling size Harper and Obeid 1967; Ackermanand Gordon 1969; Schaal 1980;
Cideciyan and Malloch 1982; Zimmermanand Weis 1983; Stanton
1984a
Leaf area Weis 1982
Cotyledonarea Weis 1982; Zimmermanand Weis 1983
Root size Ackermanand Gordon 1969; Weis 1982; Zimmermanand Weis 1983
Germination% Austinand Longden 1967; Schaal 1980; Jacobsohnand Globerson
1980
Emergence% Austin and Longden 1967; Stanton 1984a
Survival % Schaal 1980
TABLE 5
STUDIES SHOWING A POSITIVE RELATIONSHIP BETWEEN EGG
SIZE AND OFFSPRING CHARACTERISTICS
Offspring
Characteristic Source
Offspringsize
Insects Steinwascher1984
Amphibians Crump 1984
Hatchingsuccess
Insects Richards and Myers 1980
Survival rate
Fish Bagenal 1969
Growthrate
Insects Steinwascher1984
exhibiting a wide rangeof seed size allow a moreprecisedetermination of the
relationship betweenoffspring size and offspring It appearsthatseedling
fitness.
performance in a varietyof charactersincreasesin a convexmannerwithseed
size (table4). Less informationis availableon theshapesoftheserelationships in
animals(butsee table5), and moreresearchis needed.
We and othershave assumedthatthe controlover investment lies withthe
parentratherthanwiththeoffspring andthatoffspring are (genetically)identical,
apartfromtheirsize. Parent-offspring conflictoverinvestment or differences in
thegeneticqualityofyoungcouldalso lead to variation inoffspring size (Trivers
1974;Temme1986).We also assumethatorganisms arephysiologically capableof
investing thesameamountofresourcesin each youngandthatthereis no trade-
offbetweenuniform investment andothertraitsofan organism. Failuretoinclude
all of the relevantconstraints in our modelsmayhelp explainthe lack of fit
betweenobservation and theory.Ratherthanassumingthatoffspring size varia-
tionis an adaptivesolutionto variableenvironments, we suggestthatitwouldbe
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 383
moreusefulto examinehow naturalselectionhas influenced the evolutionof
parentalinvestment in responseto environmental and developmental
variability
in Charnov1982).In our
(see thediscussionof "selectionthinking"
constraints
discussionof developmental we emphasizeplantsbecause more
constraints,
informationis availableon them.
Constraintsin Plants
Pleiotropic effects.-Plantsmightbe capableofproducing moreevenlysized
seeds,buttheadvantageofdoingso couldbe offset by negativeeffectson other
traits.Plantgrowth,development, andallocationpatterns bycom-
are controlled
poundsfromfivemajorclasses of planthormonesor growthsubstances.Com-
poundsfromeach groupare thought to be presentinall cells,anddifferent
ratios
ofthesesubstancesare thought to influencethedevelopment ofmanyplanttraits
(Vanderhoef and Kosuge 1984).If thisis true,it becomesdifficultto studythe
behavior of one plant traitindependentlyof all othersbecause a change in the
hormonalbalance that affectsone traitpositively(e.g., makingseed size more
uniform) mightaffectanothertraitnegatively.
Sinnottsuggestedthatthe size of a plantorgandependson the "size of the
growingpointfromwhichit developed" (1921, p. 402). This argumentwas
generalized byGrafius(1978),whoproposedthatorgansize is correlated withthe
size of the meristematic tissuesthatproducedit and thatthereare oftensize
correlations betweenneighboring plantparts.For example,ifleafsize and seed
size are related,
allometrically seed mayfollowsimplyfromselec-
size variation
tionfordifferences correlated
in leafsize withina plant.Seed size is positively
withthe size of floralorgansin wheat(Triticumaestivum; Hamidand Grafius
1978;MilletandPinthus1984),withpodwidthinsoybeans(Glycinemax; Bravoet
al. 1980;Frankand Fehr1981),andwiththesize ofcone scalesinlodgepolepine
(Pinus contorta;McGinley,C. C. Smith,and P. F. Elliott,MS).
Positional, temporal, and environmentaleffects.-Plants may have only a
limitedabilityto control seed size variationbecause size is influencedby the
positionon theplantwherea seed develops,bythetimeofseed maturation, and
by environmental conditionsduringdevelopment (table6). In manycases, the
influences of position,timing,and environment on seed size maybe related.
Environmental conditionschangeoverthecourseofa seasonand are therefore
correlatedwithtimingeffectson seed development and withpositioneffectsif
seeds developfromdifferent partsofa plantthroughout theseason.
Thefinalsize ofa seed dependson therateandduration ofseedfilling(Daynard
et al. 1971;S. Kaplan and Koller 1974;Pinthusand Sar-Shalom1978;D. Jones
et al. 1979;Simmonsand Crookston1979;Gbikpiand Crookston1981;Gebey-
ehouet al. 1982).These in turnare affected by environmental conditionsduring
development (table7). Generally,hightemperatures increasethegrowthrateof
seedsbutdecreasetheduration Hightemperatures
ofseedfilling. andwaterstress
appear to shortenthe durationof seed fillingby hasteningthe onset of leaf
senescence.Thusfar,theeffectof theenvironment on therateand durationof
seed fillinghas been examinedonlyin crop species,and its importance as a
constrainton uniform seed investment in wildplantsis unknown.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
TABLE 6
FACTORS AFFECTING OFFSPRING SIZE
Factor Sources
PLANTS
PositionEffects
Betweenbranches Anslow1964;MaunandCavers1971b;Bean 1972;Egliet al.
1978;T. Thomaset al. 1978,1979;Clarke1979;Hendrix1979;
Hurkaand Beneweg1979;Sheldrake and Saxena 1979;
Hebblethwaite et al. 1980;JacobsohnandGloberson1980;
Waller1982;Pitelkaet al. 1983;Hardin1984;HeindlandBrun
1984;Thompson1984;Spaethand Sinclair1984;Marshallet al.
1985
Within
branches MaunandCavers1971b;Spaethand Sinclair1984
Within
fruit Anslow1964;R. J.Lambertet al. 1967;RawsonandEvans 1970;
Janzen1977b;Bremner andRawson1978;Hebblethwaite et al.
1980;Schaal 1980;Pitelkaet al. 1983;Stanton1984b;Mazeret
al. 1986
SeasonalEffects Anslow1964;Frost1971;Salisbury1976;SofferandSmith1974;
Egliet al. 1978;Hendrix1979;O'Toole 1982;Caversand Steel
1984;Marshallet al. 1985
Environmental
Effects
Temperature R. G. Lambertand Linck1958;Maunet al. 1969;Wardlaw1970;
Bean 1971;Skermanand Humphreys 1973;Fordet al. 1976;
Akpanand Bean 1977;Woodet al. 1980;Campbellet al. 1981;
Grayand Steckel1984;R. J.Joneset al. 1984
Moisturelevel D. Lambert1967;Hebblethwaite 1977;Brocklehurst et al. 1978;
Jenner1979;Campbellet al. 1981;Meckelet al. 1984;Ramseur
et al. 1984
Irradiance
level Wardlaw1970;Brocklehurst et al. 1978;Jenner 1979;
Martinez-CarrascoandThorne1979
Photoperiod Bean 1971;Cook 1975;J.ThomasandRaper1976
Nutrient
level D. Lambert1967;Bean 1971;Ene andBean 1975;Hebblethwaite
1977;Streeter1978;ParrishandBazzaz 1985
Defoliation Urieet al. 1968;MaunandCavers1971a;Bremner 1972;Reed
and Stephenson 1972;Janzen1976;Bentleyet al. 1980;
Stephenson 1980;Marshallet al. 1986
Seed predation Maunand Cavers1971a;R. Smithand Bass 1972;Bremner and
Rawson1978;Pinthus andMillet1978;I. WilliamsandFree
1979;Woronecki et al. 1980;Marshallet al. 1985
ANIMALS
SeasonalEffects
Cirripeds Barnesand Barnes1965
Cladocerans Kerfoot1974
Isopods Brodyand Lawlor1984
Insects Harvey1977;RichardsandMyers1980;Wiklund andPersson
1983;Karlssonand Wiklund1984
Fish de Ciechomski1966,1973;Hempeland Blaxter1967;G. Williams
1967;Hubbset al. 1968;Bagenal1971;Ware1977;Hislopet al.
1978;Marsh1984
Amphibians Howard1978
MaternalSize or Condition
Cirripeds Barnesand Barnes1965
Insects R. E. Joneset al. 1982
Fish de Ciechomski 1966;Hubbset al. 1968;Wootton1973;Hulataet
al. 1974
Amphibians Saltheand Duelman1973;R. Kaplanand Salthe1979;Fraser1980
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 385
TABLE 7
STUDIES SHOWING A RELATIONSHIP BETWEEN ENVIRONMENTAL CONDITIONS DURING SEED DEVELOPMENT
AND THE RATE AND DURATION OF SEED FILLING
Sources
Rate of Seed Filling
Temperature Ford et al. 1976;Sofieldet al. 1977;Egli and Wardlaw 1980;R. J.
Jones et al. 1981,1984;Donovan et al. 1983
Moisturelevel Brocklehurstet al. 1978
Irradiancelevel Brocklehurstet al. 1978;Jenner1979
Durationof Seed Filling
Temperature Marcellos and Single 1972;Ford et al. 1976;Hunteret al. 1977;Egli
and Wardlaw 1980;R. J. Joneset al. 1981,1984
Moisturelevel Brooks et al. 1982;Korte et al. 1983;Meckel et al. 1984
Seed size variationcould also arise if a plantincorrectlyassesses resource
forseed filling.Manyplantsuse the photosynthate
availability producedafter
to matureseeds (Bewleyand Black 1985)and maytherefore
seed initiation not
"know" the exact resourcepool availableforseed filling at the timeof seed
Plantsshouldattempt
initiation. to adjustthenumberofflowersand ovulesthey
makeand the fractionof thesethattheyallow to matureon the basis of the
expectedresourcepool. However,becauseofthemanyfactorsthataffect photo-
syntheticrateand resourceacquisition, plantsmayoftendevelopmoreor fewer
seeds,and hencemakesmalleror largerseeds,thanwouldbe optimal.
For example,herbivory on leaves and seed predationmaychangethebalance
betweentheexpectedresourcepool and thenumberofseedsproduced.Herbiv-
ory decreasesresourceavailability, whereasseed predationincreasesthe re-
sourcesavailableto theremaining seeds. Plantscan respondto alteredresource
levelsby changingseed numberor seed size. Defoliation causes a reductionin
seed number(McAllisterand Krober1958;Maunand Cavers 1971b;Rockwood
1973;Clarke1978;G. Thomaset al. 1978;Bentleyet al. 1980;Stephenson1980;
Marshallet al. 1985)and seed size (table6) in manyspecies.Conversely,seed
predation oftenresultsin largerseeds (table6).
Because plantsseriallyadjustinvestment to femalereproduction (Lloyd 1980),
thetiming ofeventsaffecting reproduction seed size. For example,
can influence
if plantscannotrecoverresourcesalreadyinvestedin seeds that,for some
reason,are to be aborted,these seeds will be smallerthannormal.Seed size
differencesmayresultif decisionsto abortare made at different times(Wiens
1984).
Finally,we haveassumedinourmodelsthatparentsareprovisioning offspring
froma commonresourcepool. Thismaynotalwaysbe thecase inplants,which
have a modulargrowthform.Different modules(shoots,branches)mayact as
separatephysiologicalunits(Watsonand Casper 1984).Differences in seed size
amongmodulesmay resultfromthe fact thatseparatemodulesface differ-
entenvironments and must"make theirown decisions"aboutseed provision-
ing.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
386 THE AMERICAN NATURALIST
Constraintsin Animals
The factorsinfluencingoffspring
size havenotbeenas wellstudiedforanimals
as theyhaveforplants,butsimilarpatterns exist.Offspring
size changesseason-
ally,and it is affected
by environmentalconditionsduringdevelopment and by
maternalcondition(table6).
Canalizing Selection in Variable Environments
Althoughthe Smith-Fretwell modelpredictsa singleoptimaloffspring size,
canalizingselectionforthissize is unlikelyto eliminateall size variation,
particu-
larlyin viewofthedevelopmental constraintsdiscussedabove. Furthermore, in
variableenvironments, theoptimaloffspring size mayvaryfromplacetoplaceor
fromyeartoyear,thusweakening thestrength ofcanalizingselection.As a result,
we can expectgreateroffspring size variationin variableenvironments thanin
stableenvironments, notforadaptivereasons,butbecause selectionis weaker.
Researchershave, however,oftenarguedforan adaptiveexplanationwhen
seed or eggsize variationhas beenfound.For example,Crump's(1981)studyof
eggsize infrogsoftemporary and permanent pondsis oftencitedas supportfor
the hypothesis thatoffspring size variationis an adaptiveresponseto variable
environments. Temporary pondsrepresent unpredictable environments because
theydryup at different ratesfromyearto year,whereaspermanent pondsare
stablehabitats.Crumppredictedthatfrogsof temporary ponds shouldexhibit
greatervariationin eggsize (largercoefficient ofvariation) thanpermanent-pond
breedersand thatthe distribution of egg sizes withina clutchwouldbe more
platykurtic
forfrogsof temporaryponds and moreleptokurticforfrogsof perma-
nentponds.Crumpfoundno difference betweenthetwotypesofbreedersinthe
of variationof egg size. Only two of the seven clutchesof pond
coefficient
breedersshe studiedweresignificantlyleptokurtic,and noneof theclutchesof
permanent-pond breedersdiffered fromnormal.Crumparguedthat
significantly
thesignificant
differencebetweenthesignsoftheG2 valuesfortemporary- and
permanent-pond breederssupportedtheadaptive-size-variation hypothesis.We
believethattheevidenceis notconvincing,especiallysincegreater size
offspring
can be expectedin variableenvironments
variation fornonadaptive reasons.We
should not ask simply whether offspringare more variable in heterogeneous
environments,but ratherhow muchmorevariabletheyare and whetherthe
increasedvarianceis morethancan be expectedfromrelaxedselection.
Selectionfor OffspringSize Variability
Thusfar,we have arguedthatenvironmental variability
does notnecessarily
favorvariationin offspringsize and thatthe observedvariationin the size of
youngmayresultfromconstraints on an organism'sabilitytoregulateinvestment
in itsyoungadequately.
Nonetheless,accordingto ourmodels,somecircumstances favoroffspringsize
variation.We findthatdensitydependencein offspring fitness
is requiredforthe
selectionof variableinvestment in spatiallyheterogeneous environments. In
addition,parentsmustbe capableofproducing offspringoftheappropriate size
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 387
forthehabitatsinwhichoffspringmature.Thiscouldbe accomplished bymobile
parentswho assess habitatqualityand makeoffspring of the correctsize, by
immobileparentswhocan controlthedispersaloftheiroffspring,orbyoffspring
whocan selecttheappropriatehabitatfortheirsize.
Animals are more likelyto meet these conditionsthanare plants,and animals
are thereforemorelikelyto show adaptive size variationin young.Because plants
are immobile,theymustrelyon externalvectorsto disperse theirseeds (van der
Pijl 1982). They may controlthe mechanismof seed dispersal by adjustingfruit
characteristics,but the finalrestingplace of a seed is largelycontrolledby the
movements ofthedispersing agent(animal,wind,water,etc.). It is unlikelythat
plantshave sufficient controloverthemovements oftheirdispersers to regulate
thelocationsof seeds precisely.
Some plantsmay benefitfromthe directeddispersalof seeds by ants. Ant
dispersalof Carexpedunculataseeds to rotting logs places seeds in siteswith
lowerdensitiesofinterspecific competitors (Handel1978).Otherantsreturn seeds
to theirnests,wheregermination conditionsare better(highermoistureand/or
nutrient levels) (Culverand Beattie1980;Davidsonand Morton1981;Horvitz
1981).Ifantsdispersemanyseedstothesepreferred severeintraspecific
habitats,
competition mightresultand reducea seedling'schanceofestablishment. Plants
withthissortofdispersalmight thenbe selectedtoproducesomeseedswithfood
rewardsforantdispersersand otherseeds withalternate modesofdispersal.As
faras we know,sucha dispersalsystemhas notyetbeenfound.
In animals,mobilityand the possessionof a sensorysystempermithabitat
selectionby eithertheparentor theoffspring. Femalesof manyinsectspecies
exhibitchoice in ovipositionsites (Mitchell1975;Rausher1979;Charnovand
Skinner1984;Chewand Robbins1984;Singer1984),and in thesesamespecies,
offspring fitnesshas been shownto varywithdensity.For example,survivalof
the eggs of the bean weevil (Callosobruchusmaculatus)on mung beans
(Phaseolus aureus)increaseswithbean size and decreaseswiththenumberof
eggslaid on thebean (Mitchell1975).If therelationship betweenegg size and
offspring fitnessdependson bean size and the numberof eggs per bean, the
optimaleggsize maydiffer betweenbeans.The observation thatfemaleslayeggs
on thelargerbeansfirst anddo notlaya secondeggon a beanuntilall beanshave
at least one egg suggeststhatfemalescan assess bean quality.It does not
necessarilyfollowthatfemalescan adjustoffspring size accordingto habitat
qualitybecauseofthedifference in timescale at whichoviposition decisionsare
maderelativeto theduration ofeggmaturation. In otherwords,although females
canquicklyassess habitatquality,theycannotaltereggsizeas quickly.However,
ifbeanqualityandtheoptimaloffspring size changeina predictable mannerover
theseason,femalesmightvaryeggsize seasonally.
Seasonal adjustment in offspringsize in responseto environmental variation
shouldoccur if conditionsfacedby the motherare accuratepredictors of the
circumstances youngwillencounter. This is mostlikelywhenoffspring develop
immediately and do not undergoa periodof diapause. For example,in the
terrestrialisopod Armadillium vulgare,whose offspring developdirectlyfrom
eggs,food affectsthe size of youngproducedby females(Brodyand Lawlor
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
388 THE AMERICANNATURALIST
1984).In thelab, femalesproducelargeroffspring whenfoodavailability is low;
thisis consistentwithfieldobservations ofa declineinoffspring size fromspring
to summer anda concomitant increaseinfoodresources.Similarly, eggsize inthe
cladoceranBosminalongirostris changesduringtheseason; and Kerfoot(1974)
suggested thattheshiftin eggsize representsan adaptiveresponseto changesin
predationpatterns.In orderto demonstrate thattheseshiftsare adaptive,one
wouldhave to showthattherelationship betweenoffspring fitnessand offspring
size changesin thepredictedmanneras theenvironment changes.
The use ofenvironmental cues to adjustoffspringsize wouldnotbe selected,
however,ifall offspring, no matterwhentheyweremade,maturedat thesame
time.For example,seasonalchangesin seed size wouldnotrepresent adaptive
shiftsifall seedsgerminated at thesametimethefollowing season.The germina-
tionrequirements ofseedsproducedatdifferent timesduring a seasonarenotwell
known.
SUMMARY
It has been arguedthatenvironmental favorsvariationin offspring
variability
size. We haveexaminedtheoretically theeffectsofthreetypesofenvironmental
heterogeneity on parentalinvestment: spatialvariation,spatialvariationwith
density-dependent offspring fitness,and temporal We concludedthat
variability.
environmental heterogeneity rarelyfavorstheproduction of variableoffspring.
Withspatialheterogeneity andnodensitydependenceinfitness, a singleoffspring
size is alwaysoptimal.Variableoffspring maybe favoredifdensity-dependent
fitnessloss tooffspring is largeandifparentscan controlthedispersalofoffspring
to theappropriate habitats.If environments varytemporally, thepredictedop-
timalparentalinvestment strategydependson themeasureoffitness used. If the
arithmetic mean of fitnessis used, a singleoffspring size is alwaysfavored.
Variableoffspring sizes are sometimesselectedif fitnessis measuredby the
arithmetic mean discountedforthe varianceor by the geometricmean. Our
resultsdiffer fromthoseofR. KaplanandCooper(1984)becauseofdifferences in
ourassumptions abouttherelationship amongoffspring fitness,offspring size,and
environmental quality.
Although a singleoffspring size is oftentheoptimalparentalinvestment strat-
egy accordingto our models,considerablevariationin the size of eggs/seeds/
younghas beenfoundinanimalsandplants.We suggestthatan organism's ability
tocontrolinvestment inoffspring precisely andtoreducethevariation inoffspring
size maybe constrained by suchfactorsas pleiotropy and positional,temporal,
and environmental effects duringoffspring development.
We suggestthatanimalsare morelikelyto exhibitadaptiveoffspring size
variationthanplantsbecause of theirgreaterabilityto assess environmental
conditions and to disperseoffspring to appropriatehabitats.
ACKNOWLEDGMENTS
We thankthefollowing
forhelpfuldiscussionsand comments on earlierdrafts
of thismanuscript:
M. Aide, S. Carroll,E. Charnov,V. Eckhart,S. Parkinson,
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 389
T. Philippi,D. Samson, C. Smith,S. Sutherland,J. Zimmerman,and membersof
the Ecology and Evolution JournalClub at the Universityof Utah. The figures
were preparedby K. Matz.
APPENDIX A
A parentshouldproduceoffspring smallerthanthe minimum viable size in a poor
environment (i.e., s < km) if,in so doing,its fitnessis greaterthanthatof a parent
producing offspring largerthanthissize.
First,we establishtheoptimaloffspring sizesforparentsmaking youngsmallerorlarger
thankm.Then,we determine whenthefitness parentexceedsthatofthesecond.
ofthefirst
If s < km,parentalfitnessdependsonlyon the successof youngdispersing to good
habitats: W(s) = pN(s)Fg(s). Parental fitnessis maximizedwhen S = g9,provided that
3g < km.
Ifs > km,parentalfitness
dependson thesuccessofoffspring to bothhabitat
dispersing
types:
W(s) = pN(s)Fg(s) + (1 - p)N(s)Fb(s).
Parentalfitness
is maximized
when
, a
= = m(1 + a)l/a[p + (1 p)ka]1-
Note that gmis intermediatein size between Sg and gb. Provided that Sg < km, when is
W(Sg)> W(&m) or pN(Sg)Fg(Sg)> pN(&m)Fg(&m) + (1 - p)N(Sm)Fb(Sm) ?
Substitutingforg9,gm,N(s), Fg(s), and Fb(s) andsolving,we findthatparentsproducing
whenk > A, where
smallerthankmhavehigherfitness
offspring
A = [(1 _ pl+a)lpa(1 _ p)]l/a,
providedthatg9< km.Conversely,
parentsshouldproduceyoungofintermediate
size (sm)
when k < A or when 3g > km.
APPENDIX B
The setofall two-offspring-size
strategies butwe can readilyeliminate
is clearlyinfinite,
manyofthembecausetheyneverconferhigherfitness thana single-sizestrategy.
We can disregard thefollowingtwo-sizestrategies to a one-size
as viablealternatives
strategy.
Bothoffspring largerthankm.-We haveshownthatwhenconditions favortheproduc-
tionofoffspring largerthankm,gm is thesize thatyieldsthehighest fitness-to-investment
ratioforparents.It followsthatparentalfitnesscan neverbe increased,underthese
conditions,
by producing twooffspring sizes bothlargerthankm.
Bothoffspring smallerthankm.-By the same reasoning, whenconditions favorthe
productionofoffspring smallerthankm,parental fitnessis maximizedbymaking offspring
of size Sg.
We cannotas yetruleoutthepossibility thata combination oftwosizes,onelargerthan
kmand theothersmaller,wouldoutcompete a single-size
strategy.First,letus consider
whattheoptimalsize ofthesmalleroffspring shouldbe. Becauseoffspring smallerthankm
surviveonlyinthegoodhabitat,itis clearthatthesmallersize shouldbe g9(provided that
S < km).Theoptimalsizeofthelargeroffspring is lessobvious,however, andcouldbe gm,
Sb, or someothersize.
We must then consider strategiesof the form(Qg, vsm, q), where v is a constant(v >
kml&m)and q is thefraction of parentalresourcesallocatedto smalloffspring. We then
determine thevalueofv forwhichparental fitness is maximized;we can thencomparethe
fitness ofa parentusinga two-sizestrategywiththatofa parentusinga one-sizestrategy
(eithersg or gm)
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
390 THE AMERICAN NATURALIST
W(3g, q) = + (1 - + (1 - p)N(vSm)Fb(vSm)I.
vsm, pqN(3g)Fg(sf q)[pN(vS^m)Fg(vm)
Substitutingfor gg,V&m,N(s), Fg(s), and Fb(s) and solving,we findthat
W(^g
W(s^g, s', q))=
VSm,
E
E L
F pqa
[ m(l + a)l/a(1 + a)
+ + 1-q va(l+a) -11
m(l + a)l+ l/a[p + (1 - p)kIl/a Vl+a
where E is the parentalresource level. We also findthat maximizingW(?g,vsm, q) with
respectto v is equivalentto maximizing theexpressionY = [va(l + a) - lv +a with
respectto v.
The value of v thatmaximizes Y and W(Sg,vsm,q) is v = 1. Hence, ifa parentuses a two-
itsfitness
size strategy, is highestifitproducesa combination
of small(sg)and medium-
sized (s^m)young.
We can nowask whena two-sizestrategy
is betterthana one-sizestrategy.
We findthat
W(&g,3m, q) > W(g) if and only if k < A,
whereA = [(1 _ pl+a)Ipa(1 _ p)]l/a;
W(Sg,3m, q) > W(3m) if and only if k > A.
(Ifk = A, thenall threestrategies
do equallywell.)FromAppendix
A, we knowthatW(3g)
> W(&m)when k > A. Combiningthese inequalities,we findthat
W(g)> W(?,9 g q) > W(S^M) 9
when k > A and, conversely,thatwhen k < A,
W(?g< (g, sm, q) < W(3m)-
Thus,a two-sizestrategy
neverdoes betterthana one-sizestrategy.
APPENDIX C
To characterize
densitydependenceinoffspringfitness,we assumethatoffspringfitness
declinesin proportion
to thenumberof youngpresentin a habitat.Thus,ifthereare M
of size s has fitnessFg(s)(l - cM), wherec
youngin the good habitat,an offspring
measurestheintensityof density-dependent
fitness an offspring
loss. Similarly, of size s
who sharesthebad habitatwithM' younghas fitness Fb(s)(l - cM'). The numbersof
young,M andM', arefunctions oftheparentalinvestment usedbyparentsinthe
strategy
population.
We considertwoalternative SI, parentsproducea singleoffspring
strategies: size that
maximizes parentalfitnessunderrandomdispersal(i.e., Sgor 3m);and S2, parentsproducea
combination of large(Qb) and small(sg) offspring
and attempt to dispersethemto the
appropriatehabitats.
Ifthepopulation consistsofP parentsusingstrategySI, thenumber ofyounginthegood
habitatis
M= pEPIs, (Cl)
andthenumberofyoungin thebad habitatis
M' = (1 -p) EPIs, (C2)
wherep is thefractionofgoodhabitatandE is theparentalresourceforreproduction.
ifthepopulationconsistsofP parentsusingstrategy
Alternatively, S2, thenumberof
in thetwohabitatsis
offspring
M = p [qEP(l + h)Igg + (1 - q)EP(l - h)/lb] (C3)
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 391
and
M' = (1 - p) [qEP(I - h)l/g + (1 - q)EP(I + h)/3b], (C4)
whereq is thefraction ofparentalresourcesdevotedto smalloffspring andh measuresthe
abilityofparentsto directoffspring to theappropriate habitatfortheirsize.
We arenowina positionto determine theexpectedfitnessofa rareparentusingstrategy
Si (i = 1, 2) in a populationof Sj (j = 1, 2; i #j) parentsand theexpectedfitness of Si
parentsin an Si population (i = 1, 2). In eithercase thefitnessofa parentis simply
NgFg(l - cM) + NbFb(l - cM'),
whereNg and Nb are the numberof a parent'soffspring in thegood and bad habitats,
respectively, and whereM and M' are definedas in equations(CI) and (C2) or (C3) and
(C4), depending on whetherthepopulation consistsofSI or S2 parents.
Forexample,theexpectedfitness ofa parentproducingmedium-sized offspring(strategy
SI withs = sm)in a populationof like parentsis
E(S1, S1) = p(EIs^m)Fg(?m) (1 - cpEPISm) + (1 - p)(EISm)Fb(Sm) [1 - c(1 - p)EPISm].
ofa rareS2 parentin a population
The expectedfitness producing youngis
medium-sized
E(S2, S1) = pE [q( + h)Fg(sg) ( cpEP (I - q)( -h)Fg(9b) ( c
L Sg sm ISb s
-
+ (1 -p)E [q( h)Fb(Sg) (1 c(l -p)EP)
+ (I-q)(I + h)Fb(Sb) (1 c(l p)EP)1
Sb 3m
LITERATURE CITED
Ackerman, R. F., and J. R. Gorman.1969.Effectofseed weighton thesize oflodgepolepineand
whitesprucecontainer-planting stock.PulpPaperMag. Can. 70:167-169.
Akpan,E. E. J.,and E. W. Bean. 1977.The effect oftemperature uponseed development in three
speciesofforagegrasses.Ann.Bot. (Lond.) 41:689-695.
Anslow,R. C. 1964.Seed formation inperennialryegrass.
II. Maturationofseed.J.Br. Grassl.Soc.
19:349-357.
Austin,R. B., andP. C. Longden.1967.Someeffects ofseedsize andmaturity on theyieldofcarrot
crops.J.Hortic.Sci. 42:339-353.
Autenson, H. M., andP. D. Walton.1970.Relationship betweeninitialseedweight andmature plant
characters in springwheat.Can. J.PlantSci. 50:53-58.
Ayers,G. S., V. F. Wert,and S. K. Ries. 1976.The relationshipofprotein fractions
andindividual
proteinto seedlingvigorin wheat.Ann.Bot. (Lond.) 40:563-570.
Bagenal,T. B. 1969.Relationship betweeneggsize andfrysurvival inbrowntrout, Salmotrutta L. J.
FishBiol. 1:349-353.
1971.Theinterrelation ofthesize offisheggs,thedateofspawning, andpopulation cycle.J.
Fish Biol. 3:207-219.
Barnes,H., andM. Barnes.1965.Egg size,naupliussize,andtheirvariation withlocal,geographic,
and specificfactorsin somecommoncirripeds. J.Anim.Ecol. 34:391-402.
Bean, E. W. 1971.Temperature effectsupon inflorescenceand seed development in tall fescue
(Festuca arundinacea). Ann. Bot. (Lond.) 35:891-897.
1972.Seed quality:itsvariation,
control
andimportance inbreeding
andvarietalassessment.
WelshPlantBreed.Stn.(Aberystwyth) Rep. 1972:194-208.
Bentley,
S., J.B. Whittaker,
andA. J.Malloch.1980.Fieldexperimentsontheeffectsofgrazing
bya
chrysomelidbeetle(Gastrophysaviridula)on seed productionand qualityin Rumexob-
tusifoliaand R. crispus. J. Ecol. 68:671-674.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
392 THE AMERICAN NATURALIST
Beveridge, J. L., and C. P. Wilsie.1959.Influence of depthof planting,
seed size, and varietyon
emergence and seedlingvigorin alfalfa.Agron.J.51:731-734.
Bewley,D. J.,andM. Black. 1985.Seeds: physiology ofdevelopment andgermination. Plenum, New
York.
Black, J. N. 1956.The influence of seed size and depthof sowingon pre-emergence and early
vegetative growth. Aust.J.Agric.Res. 7:98-109.
Boyd,W. J. R., A. G. Gordon,and L. J. La Croix. 1971.Seed size, germination resistanceand
seedlingvigorin barley.Can. J.PlantSci. 51:93-99.
Bravo,J.A., W. R. Fehr,andS. Rodriguez de Cianzio.1980.Use ofpodwidth forindirect of
selection
seed weightin soybeans.CropSci. 20:507-510.
Bremner, P. M. 1972.Accumulation ofdrymatter andnitrogen bygrainsindifferent positionsofthe
wheatear as influenced by shadinganddefoliation. Aust.J.Biol. Sci. 25:657-668.
Bremner, P. M., andH. M. Rawson.1978.Theweights grainsofthewheatearinrelation
ofindividual
to theirgrowth thesupplyofassimilates,
potential, andinteraction betweengrains.Aust.J.
PlantPhysiol.5:61-72.
Brocklehurst, P. A., J.P. Moss,andW. W. Williams.1978.Effects ofirradiance andwatersupplyon
graindevelopment in wheat.Ann.Appl.Biol. 90:265-276.
Brody,M. S., and L. R. Lawlor.1984.Adaptivevariation in offspring
size intheterrestrialisopod
Armadilliumvulgare. Oecologia (Berl.) 61:55-59.
Brooks,A., C. F. Jenner,andD. Aspinall.1982.Effects ofwaterdeficit onendosperm starchgranules
andon grainphysiology ofwheatand barley.Aust.J.PlantPhysiol.9:423-436.
Bulisani,E. A., and R. L. Warner.1980.Seed proteinand nitrogen effects uponseedlingvigorin
wheat.Agron.J.72:657-662.
Campbell,C. A., H. R. Davidson,and G. E. Winkleman. 1981.Effectof nitrogen, temperature,
growthstageand durationof moisture stresson yieldcomponents and proteincontentof
Manitouspringwheat.Can. J.PlantSci. 61:549-563.
Capinera,J.L. 1979.Qualitative variationinplantsandinsects:effect ofpropagule sizeon ecological
Am.Nat. 114:350-361.
plasticity.
Carleton,A. E., andC. S. Cooper.1972.Seed sizeeffects uponseedling vigorofthreeforagelegumes.
CropSci. 12:183-186.
Cavers,P. B., andM. G. Steel.1984.Patterns ofchangeinseedweights overtimeonindividual plants.
Am.Nat. 124:324-335.
Charnov,E. L. 1982.The theoryofsex allocation.Princeton University
Press,Princeton, N.J.
Charnov,E. L., and S. W. Skinner.1984.Evolutionof hostselectionand clutchsize in parasitoid
wasps.Fla. Entomol.67:5-21.
Chew,F. S., andR. K. Robbins.1984.Egg-laying in butterflies.
Symp.R. Entomol.Soc. Lond. 11:
65-79.
Cideciyan,M. A., and A. J. C. Malloch.1982.Effectof seed size on thegermination, growth, and
competitive abilityofRumexcrispusandR. obtusifolia. J.Ecol. 70:227-232.
Clarke,J.M. 1978.Theeffect ofleafremovalon yieldandyieldcomponents ofBrassicanapus.Can.
J.PlantSci. 58:1103-1105.
1979.Intra-plant variationin numbers of seeds perpod and seed weightinBrassicanapus
'tower.'Can. J.PlantSci. 59:959-962.
Cohen,D. 1966.Optimizing reproductionina randomly varying environment. J.Theor.Biol. 12:119-
129.
Cook,R. E. 1975.Thephotoinductive controlofseedweight inChenopodium rubrum L. Am.J.Bot.
62:427-431.
Cope,W. A. 1966.Growth rateandyieldinSericealespedezainrelation to seedsizeandoutcrossing.
CropSci. 6:566-568.
Crump,M. L. 1981.Variationin propagulesize as a function ofenvironmental uncertainty fortree
frogs.Am.Nat. 117:724-737.
1984.Intraclutch inHylacrucifer
eggsize variability (Anura:Hylidae).Copeia1984:302-308.
Culver,D. C., andA. J.Beattie.1980.ThefateofViolaseedsdispersed byants.Am.J.Bot.67:710-
714.
Davidson,D. W., and S. R. Morton.1981.Myrmechory in someplants(F. Chenopodiaceae) ofthe
Australian aridzone. Oecologia(Berl.)50:357-366.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 393
Daynard,T. B., J. W. Tanner,and W. G. Duncan.1971.Durationofthegrainfilling periodand its
relationto grainyieldin corn,Zea maysL. CropSci. 11:45-48.
de Ciechomski, J. D. 1966.Development of thelarvaeand variations in thesize oftheeggsofthe
Argentine anchovy,EngraulisanchoitaHubbsandMarini.J.Cons. Cons. Int.Explor.Mer
30:281-290.
1973.The size ofeggsoftheArgentine anchovyEngraulisanchoita(Hubbsand Marini),in
relationto theseasonoftheyearandarea ofspawning. J.FishBiol. 5:393-398.
Demirbicakmak, A., M. L. Kaufmann, and L. P. V. Johnson.1963.The influence of seed size and
seedingrateon yieldand yieldcomponents ofbarley.Can. J.PlantSci. 43:330-337.
Dolan,R. W. 1984.The effect ofseed size andmaternal sourceon individual size ina population of
Ludwigialeptocarpa(Onagraceae).Am.J. Bot. 71:1302-1307.
Donovan,G. R., J. W. Lee, J. J. Longhurst, and P. Martin.1983.Effectof temperature on grain
growth andprotein accumulation incultured wheatears.Aust.J.PlantPhysiol.10:445-450.
Egli, D. B., and I. F. Wardlaw.1980.Temperature responsesof seed growthcharacteristics of
soybeans.Agron.J.72:560-564.
Egli,D. B., J.E. Leggett,andJ.M. Wood. 1978.Influence ofsoybeanseed size andpositionon the
rateandduration offilling.
Agron.J.70:127-130.
Ene, B. N., and E. W. Bean. 1975.Variation in seed qualitybetweencertified seedlotsofperennial
ryegrass andtheirrelationshipto nitrogen supplyand moisture statusduringseed develop-
ment.J. Br. Grassl.Soc. 30:195-199.
Evans,L. E., and G. M. Bhatt.1977.Influence of seed size, proteincontent,and cultivaror early
seedlingvigorin wheat.Can. J.PlantSci. 57:929-935.
Ford,M. A., I. Pearman,andG. N. Thorne.1976.Effects ofvariationineartemperature on growth
and yieldofspringwheat.Ann.Appl.Biol. 82:317-333.
Fowells,H. A. 1953.Theeffect ofseedandstocksizeson survival andearlygrowth ofponderosaand
Jeffreypine.J.For. 51:504-507.
Frank,S. J.,andW. H. Fehr.1981.Associations amongpoddimensions andseedweight insoybeans.
Crop Sci. 21:547-550.
Fraser,D. F. 1980.On theenvironmental controlofoocytematuration ina plethodontidsalamander.
Oecologia(Berl.)46:302-307.
Frost,R. A. 1971.Aspectsof the comparative biologyof threeweedyspeciesof Amaranthus in
southwestern Ontario.Ph.D. diss. University ofOntario,Ottawa.
Gbikpi,P. J.,andR. K. Crookston.1981.Effectofflowering dateon accumulation ofdrymatter and
proteinin soybeanseeds. CropSci. 21:652-655.
Gebeyehou, G., D. R. Knott,andR. J.Baker.1982.Rateandduration inDurumwheat
ofgrainfilling
cultivars.CropSci. 22:337-340.
Glewen,K. L., andK. P. Vogel.1984.Partitioning thegeneticvariabilityforseedlinggrowth in sand
bluestemintoseed size and seedlingvigorcomponents. CropSci. 24:137-141.
Grafius,J. E. 1978.Multiplecharactersand correlated response.CropSci. 18:931-934.
Gray,D., and J. R. A. Steckel. 1984. Viabilityof onion(Alliumcepa) seed as influenced by
temperature duringseed growth. Ann.Appl.Biol. 104:375-382.
Gross,K. L. 1984.Effectofseed size andgrowth formon seedlingestablishment ofsixmonocarpic
perennials. J.Ecol. 72:369-387.
Hamid,Z. A., andJ.E. Grafius.1978.Developmental allometryanditsimplication to grainyieldin
barley.CropSci. 18:83-86.
Handel,S. N. 1978.The competitive relationship of threewoodlandsedgesand itsbearingon the
evolutionofant-dispersal ofCarexpedunculata.Evolution32:151-163.
Hardin,E. D. 1984.Variationin seed weight,numberper capsule,and germination in Populus
deltoidesBartr.treesin southeastern Ohio.Am. Midl.Nat. 112:29-34.
Harper,J.L., andM. Obeid.1967.Influence ofseedsizeanddepthofsowingontheestablishment and
growth ofvarietiesoffiberandoil seed flax.CropSci. 7:527-532.
Harvey,G. T. 1977.Meanweight andrearing performance ofsuccessiveeggclustersofeasternspruce
budworm (Lepidoptera: Torticidae).Can. Entomol.109:487-496.
Hebblethwaite, P. D. 1977.Irrigation
andnitrogen studiesinS.23ryegrass grownforseeds.I. Growth,
development, seed yieldcomponents, and seed yield.J.Agric.Sci. 88:605-614.
ed. 1980.Seed production. Butterworth, London.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
394 THE AMERICAN NATURALIST
Hebblethwaite, P. D., D. Wright, and A. Noble. 1980.Some physiologicalaspectsof seed yieldin
Loliumperenne(perennial ryegrass).
Pages71-90 inHebblethwaite 1980.
Heindl,J. C., and W. A. Brun. 1984.Patternsof reproductive abscission,seed yield,and yield
components in soybeans.CropSci. 24:542-545.
Hempel,G., andJ.H. S. Blaxter.1967.Eggweight inAtlanticherring
(Clupeaharengus L.). J.Cons.
Cons. Int.Explor.Mer31:170-195.
Hendrix,S. D. 1979.Compensatory reproductionin a biennialherbfollowing insectdefoliation.
Oecologia(Berl.)42:107-118.
1984.Variation in seed weightanditseffects
on germinationinPastinacasativaL. (Umbel-
liferae).Am.J. Bot. 71:795-802.
Hislop,J.R. G., A. P. Robb,andJ.A. Gauld.1978.Observations oneffectsoffeeding levelongrowth
and reproductionin haddock, Melanogrammusaegelfinus(L.). J. Fish Biol. 13:85-98.
Horvitz,C. C. 1981.Analysisof how ant behaviorsaffectgermination
in a tropicalmyrmechore
Calatheamicrocephala(P&E) Koerricke(Marantaceae):microsite andarilremoval
selection
by Neotropicalants, Odontomachus,Pachycondyla,and Solenopsis (Formicidae). Oecologia
(Berl.) 51:47-52.
Howard,R. D. 1978.The influence ofmale-defended oviposition siteson earlyembryomortality in
bullfrogs.Ecology59:789-798.
Howe,H. F., andW. M. Richter.1982.Effectofseed size on seedlingsize in Virolasurinamensis: a
within and betweentreeanalysis.Oecologia(Berl.)53:347-351.
Hubbs,C., M. M. Stevenson,and A. E. Peden. 1968.Fecundity and eggsize in twocentralTexas
darterpopulations. Southwest. Nat. 13:301-324.
Hulata,G., R. Moav,andG. Wollfarth. 1974.Therelationship ofgonadandeggsizetoweight andage
in theEuropeanand Chineseracesofthecommoncarp,Cyprinus carpioL. J. Fish Biol.
6:745-758.
Hunter,R. B., M. Tollenaar,and C. M. Breuer.1977.Effectsof photoperiod and temperature on
vegetative andreproductive growth ofa maize(Zea mays)hybrid. Can.J.PlantSci. 57:1127-
1133.
Hurka,H., and M. Benneweg.1979.Patterns of seed size variation in populationsof thecommon
weedCapsellabursa-pastoris (Brassicaceae).Biol. Zentralbl. 98:699-709.
Jacobsohn, R., andD. Globerson.1980.Daucus carota(carrot)seedquality.I. Effects ofseedsizeon
germination andplantgrowth undersubtropical conditions.II. Theimportance oftheprimary
umbelin carrotseed production. Pages637-646inHebblethwaite 1980.
Janzen,D. H. 1976.Effectof defoliation on fruit-bearing branchesof the Kentuckycoffeetree,
Gymnocladus dioicus(Leguminosae).Am.Midl.Nat. 95:474-478.
1977a.Promising direction ofstudyintropical plant-animalinteractions.Ann.Mo. Bot.Gard.
61:706-736.
1977b.Variation inseedsizewithin a cropofa CostaRicanMicunaandreana(Leguminosae).
Am.J.Bot. 64:347-349.
Jenner,C. F. 1979.Grain-fillinginwheatplantsshadedforbriefperiodsafteranthesis.Aust.J.Plant
Physiol.6:629-641.
Jones,D. B., M. L. Peterson, andS. Geng.1979.Association betweengrainfilling rate,duration and
yieldcomponents in rice.CropSci. 19:641-644.
Jones,R. E., J.R. Hart,andG. D. Bull. 1982.Temperature, size,andeggproduction inthecabbage
butterfly,Pierisrapae L. Aust.J.Zool. 30:223-232.
Jones,R. J., B. G. Gengbach,and V. B. Cardwell.1981.Temperature effectson in vitrokernel
development ofmaize.CropSci. 21:761-766.
Jones,R. J., S. Quattar,and R. K. Crookston.1984.Thermalenvironment duringendosperm cell
divisionandgrainfilling in maize:effects on kernelgrowth anddevelopment in vitro.Crop
Sci. 24:133-137.
Kalton,R. R., andB. R. Christie.1957.Seed size anditsrelationship withotheragronomic character-
isticsin bromegrass.Rep. WesternGrassBreeders21:7-12.
Kaplan,R. H. 1980.The implications of ovumsize variability foroffspring fitnessand clutchsize
withinseveralpopulations ofsalamanders (Ambystoma). Evolution34:51-64.
Kaplan,R. H., and W. S. Cooper. 1984.The evolutionofdevelopmental in reproductive
plasticity
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 395
characteristics:an application ofthe"adaptivecoinflipping" principle. Am. Nat. 123:393-
410.
Kaplan,R. H., and S. N. Salthe.1979.The allometry ofreproduction: an empirical viewin salaman-
ders.Am. Nat. 113:671-689.
Kaplan,S. L., andH. R. Koller.1974.Variation amongsoybeancultivars in seedgrowth rateduring
thelinearphaseofseed growth. CropSci. 14:613-614.
Karlsson,B., andC. Wiklund.1984.Eggweight variation
andlackofcorrelation betweeneggweight
and offspring fitness in thewallbrownbutterfly Lasiommatamegera.Oikos43:376-385.
Kaufmann, M. L., and A. D. McFadden.1960.The competitive interaction betweenbarleyplants
grownfromlargeand smallseeds. Can. J.PlantSci. 40:623-629.
Kerfoot,W. C. 1974.Egg size cycleofa cladoceran.Ecology55:1259-1270.
Kneebone,W. R. 1955.Seedlingvigorin nativegrasses.Rep. Western GrassBreeders19:14-18.
Kneebone,W. R., and C. L. Cremer.1955.The relationship of seed size to seedlingvigorin some
nativegrassspecies.Agron.J.47:472-477.
Korte,L. L., J.E. Specht,J.H. Williams, andR. C. Sorensen.1983.Irrigation ofsoybeangenotypes
duringreproductive ontogeny. II. Yieldcomponent responses.CropSci. 23:528-533.
Lacey,E. P., L. Real, J.Antonovics, and D. G. Heckel. 1983.Variancemodelsin thestudyoflife
histories. Am.Nat. 122:114-131.
Lambert,D. A. 1967.The effectofnitrogen and irrigation
on timothy (Phleumpratense)grownfor
production of seed. II. Reproductive growth and yieldofseed. J.Agric.Sci. 69:231-239.
Lambert,R. G., and A. J. Linck. 1958.Effectofhightemperature on yieldofpeas. PlantPhysiol.
33:347-350.
Lambert, R. J.,D. E. Alexander, andR. C. Rodgers.1967.Effectofkernelpositionon oil content in
corn(Zea maysL.). CropSci. 7:143-145.
Langdon,0. G. 1958.Coneandseed size ofsouthFloridaslashpineandtheireffects on seedlingsize
and survival.J.For. 56:122-127.
Lewis,E. J.,and J. A. Garcia. 1979.The effectof seed weightand coleptiletillerdevelopment on
seedlingvigorin tallfescue,Festucaarundinacea.Euphytica 28:393-402.
Lloyd,D. G. 1980.Sexual strategies in plants.I. An hypothesis of serialadjustment of maternal
investment duringone reproductive session.New Phytol.86:69-79.
Lowe, L. B., and S. K. Ries. 1973.Endosperm proteinofwheatseed as a determinant of seedling
growth. PlantPhysiol.(Bethesda)51:57-60.
Malik,B. S., and J. S. Kanwar.1969.Effectof seed size and stageof harvestof carrotseed on
germination, growth, and yieldofcarrot.IndianJ.Agric.Sci. 39:603-610.
Marcellos,H., and W. V. Single.1972.Influence ofcultivartemperature and photoperiod on post-
flowering development in wheat.Aust.J.Agric.Res. 23:533-540.
Marsh,E. 1984.Egg size variationin centralTexas populations ofEtheostomaspectabile(Pisces:
Perchidae).Copeia 1984:291-301.
Marshall,D. L. 1986.Effectofseed size on seedlingsuccessinthreespeciesofSesbania(Fabaceae).
Am.J. Bot. 73:457-464.
Marshall,D. L., N. L. Fowler,andD. A. Levin. 1985.Plasticity in yieldcomponents in responseto
fruitpredation and date offlowerinitiation in threespeciesofSesbania(Leguminosae). J.
Ecol. 73:71-81.
Marshall,D. L., D. A. Levin,and N. L. Fowler.1986.Plasticity ofyieldcomponents in responseto
stress in Sesbania macrocarpa and Sesbania vesicaria (Leguminosae). Am. Nat. 127:508-
521.
R., andG. N. Thorne.1979.Physiological
Martinez-Carrasco, factors grainsize inwheat.J.
limiting
Exp. Bot. 30:669-679.
Maun,M. A., andP. B. Cavers.197la. Seedproductionanddormancy inRumexcrispus.I. Theeffect
ofremovalofcaulineleavesat anthesis.Can. J.Bot. 49:1123-1130.
1971b. Seed production
anddormancy inRumexcrispus.II. Theeffectsofremovalofvarious
proportionsofflowersat anthesis.Can. J.Bot. 49:1841-1848.
Maun,M. A., C. L. Canode,andI. D. Teare. 1969.Influenceoftemperatureduringanthesison seed
setinPoa pratensisL. CropSci. 9:210-212.
MaynardSmith,J.1982.Evolutionandthetheory ofgames.Cambridge Press,Cambridge.
University
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
396 THE AMERICAN NATURALIST
Mazer,S. J.,A. A. Snow,andM. L. Stanton.1986.Fertilization andparental
dynamics effectsupon
fruitdevelopmentinRaphanusraphanistrum: consequencesforseed size variation.
Am.J.
Bot. 73:500-511.
D. F., and0. A. Krober.1958.Responseofsoybeanstoleafandpodremoval.Agron.J.
McAllister,
50:674-676.
Meckel,L. B., D. B. Egli,R. E. Phillips,D. Radcliffe, andJ. E. Leggett.1984.Effectof moisture
stresson seed growth in soybeans.Agron.J.76:647-650.
Millet,E., and M. J.Pinthus.1984.Effectofremoving floralorgans,lightpenetration, andphysical
constraint on thedevelopment ofwheatgrains.Ann.Bot. (Lond.) 53:251-269.
Mitchell, R. 1975.The evolution ofovipositiontacticsinthebeanweevil,Callosobruchus maculatus
(F.). Ecology56:696-702.
Morse,D. H., and J. Schmitt.1985.Propagulesize, dispersalability,and seedlingperformance in
Asclepiassyrica.Oecologia(Berl.)67:372-379.
Nelson,D. M., and C. D. Johnson.1983. Stabilizingselectionon seed size in Astragalus(Le-
guminosae) duetodifferentialpredation anddifferential germination. J.Kans. Entomol.Soc.
56:169-174.
Oexemann,S. W. 1941.Relationof seed weightsto vegetative growth, and yieldin
differentiation,
plants.Am.J.Bot. 29:72-81.
O'Toole,J.J.1982.Seed banksofPanicummiliaceum L. inthreecrops.Master'sthesis.University of
WesternOntario,London.
Parrish,J. A. D., and F. A. Bazzaz. 1985.Nutrient contentofAbutilontheophrasti seeds and the
competitive abilityoftheresultingplants.Oecologia(Berl.)65:247-251.
Peace, R. D. 1953.Heritableand environmental relationships of seed weightand seedlingvigorin
smoothbromegrass, Bromusinermis. Rep. Western GrassBreeders7:36-38.
Peacock,H. A., and B. S. Hawkins.1970.Effectof seed sourceon seedlingvigor,yield,and lint
characteristicsofuplandcotton,Gossypium hirsutum L. CropSci. 10:667-670.
Pinthus, M. J.,and E. Millet.1978.Interactions amongnumber of spikelets,number ofgrains,and
grainweightin thespikesofwheat(Triticum aestivum).Ann.Bot. (Lond.)42:839-848.
Pinthus, M. J.,and Y. Sar-Shalom.1978.Drymatter accumulation in thegrainsofwheat(Triticum
aestivumL.) cultivars in grainweight.Ann.Bot. (Lond.) 42:469-471.
differing
Pitelka,L. F., M. E. Thayer,andS. B. Hansen.1983.Variation inacheneweight inAsteracuminatus.
Can. J.Bot. 61:1415-1420.
Ramseur,E. L., V. L. Quisenberry, S. U. Wallace,and J. H. Palmer.1984.Yield and yield
components of"Braxton"soybeansas influenced byirrigation andintrarow spacing.Agron.
J.76:442-446.
Rausher,M. D. 1979.Egg recognition: itsadvantageto a butterfly. Anim.Behav.27:1034-1040.
Rawson,H. M., andL. T. Evans. 1970.The pattern ofgraingrowth within theearofwheat.Aust.J.
Biol. Sci. 23:753-764.
Reed,F. C., andS. N. Stephenson. 1972.Factorsaffecting seednumber andsizeinburdock, Arctium
minusSchk.Mich.Acad. 4:448-455.
Richards, L. J.,andJ.H. Myers.1980.Maternal influences on sizeandemergence timeinthecinnabar
moth.Can. J.Zool. 58:1452-1457.
Ries,S. K., andE. H. Everson.1973.Protein content andseedsizerelationships withseedlingvigorof
wheatcultivars. Agron.J.65:884-886.
Righter, F. I. 1945.Pinus:therelationship of seed size and seedlingsize to inherent vigor.J. For.
43:131-137.
Rockwood,L. L. 1973.The effect ofdefoliationon seed production ofsix CostaRicantreespecies.
Ecology54:1363-1368.
Rogler,G. A. 1954.Seed size and seedlingvigorin crestedwheatgrass. Agron.J.46:216-220.
Salisbury, E. J. 1976.Exceptional fruitfulness
anditsbiologicalsignificance. Proc.R. Soc. Lond. B,
Biol. Sci. 193:455-460.
Salthe,S. N., andW. E. Duellman.1973.Quantitative constraints associatedwithreproductive mode
inanurans.Pages229-249inJ.L. Vial,ed. Evolutionary biologyoftheanurans.University
ofMissouriPress,Columbia.
Schaal,B. A. 1980.Reproductive capacityandseedsize inLupinustexensis. Am.J.Bot.67:703-709.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
PARENTAL INVESTMENT IN VARIABLE ENVIRONMENTS 397
Sheldrake, A. R., andN. P. Saxena. 1979.Comparisons ofearlierandlaterformed podsofchickpeas
(CicerarietumL.). Ann.Bot. (Lond.) 43:467-473.
Simmons, S. R., andR. K. Crookston. 1979.Rateandduration ofgrowth ofkernels formed at specific
florets in spikeletsofspringwheat.CropSci. 19:690-693.
Singer,M. C. 1984.Butterfly-hostplant hostquality,adultchoice,andlarvalsuccess.
relationships:
Symp.R. Entomol.Soc. Lond. 11:81-88.
Sinnott,E. W. 1921.The relationship betweenbodysize andorgansize inplants.Am.Nat. 55:385-
403.
Skerman,R. H., and L. R. Humphreys.1973. Effectof temperature duringflowering on seed
formation ofSylosanthes humilis.Aust.J.Agric.Res. 24:317-324.
Smith,C. C., and S. D. Fretwell.1974.The optimalbalancebetweensize and number ofoffspring.
Am. Nat. 108:499-506.
Smith,0. E., N. C. Welch,andT. M. Little.1973.Studieson lettuceseed quality.I. Effectofseed
size andweighton vigor.J.Am. Soc. Hortic.Sci. 98:529-533.
Smith,R. H., andM. H. Bass. 1972.Relationship podremovalto soybeanyield.J.Econ.
ofartificial
Entomol.65:606-608.
H., and0. E. Smith.1974.Studiesonlettuceseedquality.III. Relationship
Soffer, betweenflowering
pattern, seed yield,and seed quality.J.Am. Soc. Hortic.Sci. 99:114-117.
Sofield,I., L. T. Evans, M. G. Cook, and I. F. Wardlaw.1977.Factorsinfluencing therateand
duration ofgrainfillingin wheat.Aust.J.PlantPhysiol.4:785-797.
Spaeth,S. C., andT. R. Sinclair.1984.Soybeanseedgrowth. II. Individualseedmassandcomponent
compensation. Agron.J.76:128-133.
Spurr,S. H. 1944.Effectof seed weightand seed originon theearlydevelopment ofeasternwhite
pine.J.ArnoldArbor.Harv.Univ.25:467-480.
Stamp,N. E., and J. R. Lucas. 1983.Ecologicalcorrelates of explosiveseed dispersal.Oecologia
(Berl.)59:272-278.
Stanton, M. L. 1984a.Seed sizevariation inwildradish:effectofseedsizeoncomponents ofseedling
andadultfitness. Ecology65:1105-1112.
1984b.Developmental andgeneticsourcesofseedweight inRaphanusraphanistrum
variation
L. (Brassicaceae).Am.J. Bot. 71:1090-1098.
1985.Seed sizeandemergence timewithin a standofwildradish(Raphanusraphanistrum L.):
theestablishment ofa fitnesshierarchy.Oecologia(Berl.)67:524-531.
Steinwascher, K. 1984.Egg size variationin Aedes aegypti:relationship to bodysize and other
variables.Am.Midl.Nat. 112:76-84.
Stephenson, A. G. 1980.Fruitset, herbivory, fruitreduction, and thefruiting of Catalpa
strategy
speciosa(Bignoniaceae).Ecology61:57-64.
Stickler,F. C., andC. E. Wasson.1963.Emergence andseedling vigorofbirdsfoot as affected
trefoil
byplanting depth,seed size, andvariety.Agron.J.55:78.
Streeter,J.C. 1978.Effects ofN starvation ofsoybeanplantsatvariousstagesofgrowth on seedyield
and N concentration ofplantpartsat maturity. Agron.J.70:74-76.
Temme,D. H. 1986.Seed size variability: a consequenceofvariablegeneticqualityamongoffspring?
Evolution40:414-417.
Thomas,G. D., C. M. Ingroffo, D. B. Smith,andC. E. Morgan.1978.Effects ofsingleandsequential
defoliationson yieldand qualityofsoybeans.J.Econ. Entomol.71:871-874.
Thomas,J.F., andC. D. Raper,Jr.1976.Photoperiodic control ofseedfillingin soybeans.CropSci.
16:667-672.
Thomas,R. L. 1966.The influence of seed weighton seedlingvigorin Loliumperenne.Ann.Bot.
(Lond.) 30:111-121.
Thomas,T. H., D. Gray,and N. L. Biddington. 1978.The influence ofpositionoftheseed on the
mother planton seed and seedlingperformance. ActaHortic.(Peking)83:57-66.
Thomas,T. H., N. L. Biddington, and D. F. O'Toole. 1979.Relationship betweenpositionon the
parentplantand dormancy characteristics ofthreecultivars ofcelery(Apiumgraveolens).
Physiol.Plant.45:492-496.
Thompson, J. N. 1984.Variationamongindividual seed massesin Lomatiumgrayi(Umbelliferae)
undercontrolled conditions: magnitude andpartitioning ofthevariance.Ecology65:626-631.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
398 THE AMERICAN NATURALIST
Trivers,R. L. 1974.Parent-offspring conflict.
Am.Zool. 14:249-264.
Twamley, B. E. 1967.Seed size andseedlingvigorinbirdsfoot trefoil.
Can. J.PlantSci. 47:603-609.
Urie,A. L., L. N. Leininger, andD. E. Zimmer.1968.Effect ofdegreeandtimeofdefoliation onyield
and relatedattributes ofsafflower.CropSci. 8:747-750.
Vanderhoef, L. N., and T. Kosuge. 1984.The molecular biologyofplanthormone action:research
directions. Waverly, Baltimore,Md.
vanderPijl,L. 1982.Principles ofdispersalin higherplants.Springer-Verlag,Berlin.
Venable,D. L., andL. Lawlor.1980.Delayedgermination anddispersalindesertannuals:escapein
space andtime.Oecologia(Berl.)46:272-282.
Waller,D. M. 1982.Factorsinfluencing seed weightinImpatienscapensis(Balsaminaceae).Am.J.
Bot. 69:1470-1475.
Wardlaw, I. F. 1970.Theearlystageofgraindevelopment inwheat:responsetolightandtemperature
in a singlevariety.Aust.J.Biol. Sci. 23:765-774.
Ware,D. M. 1977.Spawning timeandeggsizeofAtlantic mackerel, Scomberscombrus, inrelationto
plankton. J.Fish.Res. BoardCan. 34:2308-2315.
Watson,M. A., andB. B. Casper.1984.Morphogenetic constraints
onpatterns ofcarbondistribution
in plants.Annu.Rev. Ecol. Syst.15:233-258.
Weis, I. M. 1982.The effectsof propagulesize on germination and seedlinggrowthin Mirabilis
hirsuta.Can. J. Bot. 60:1868-1874.
Wiens,D. 1984.Ovule survivorship, broodsize, lifehistory,breedingsystems,and reproductive
successin plants.Oecologia(Berl.)64:47-53.
Wiklund, C., and A. Persson.1983.Fecundity, andtherelation ofeggweightvariation to offspring
fitness in thespeckledwoodbutterfly, Parargeaegeria,or whydon'tbutterfly femaleslay
moreeggs?Oikos40:53-63.
Williams, G. C. 1967.Identificationandseasonalsizechangesofeggsofthelabridfishes,Tautogolab-
rusadspersusand Tautogaonitis,ofLongIslandSound.Copeia 1967:452-453.
Williams, I. H., andJ.B. Free. 1979.Compensation ofoil-seedrape(BrassicanapusL.) plantsafter
damageto theirbudsandpods. J.Agric.Sci. 92:53-59.
Williams, W. A. 1967.Seedlinggrowthof a hypogeallegume,Viciadasycarpa,in relationto seed
weight.CropSci. 7:163-165.
Wood,D. W., R. K. Scott,andP. C. Longden.1980.Theeffect ofmother-plant temperature on seed
qualityinBeta vulgarisL. (sugarbeet).Pages257-270inHebblethwaite 1980.
Wootton, R. J. 1973.The effectof size offoodrationon eggproduction in thefemalethree-spined
stickleback, GasterosteusaculeatusL. J.FishBiol. 5:89-96.
Woronecki, P. P., R. A. Stehn,andR. A. Dolbeer.1980.Compensatory responseofmaturing corn
kernelsfollowing simulateddamagebybirds.J.Appl.Ecol. 17:737-746.
Wulff,R. 1973.Intra-populational variationinthegermination ofseedsofHyptissuaveolens.Ecology
54:646-649.
Zimmerman, J.K., andI. M. Weis.1983.Fruitsizevariation anditseffect
ongermination andseedling
growth inXanthium strumarium. Can. J.Bot. 61:2309-2315.
This content downloaded from 132.236.27.111 on Sun, 9 Mar 2014 18:02:25 PM
All use subject to JSTOR Terms and Conditions
View publication stats
You might also like
- Biochar Application: Essential Soil Microbial EcologyFrom EverandBiochar Application: Essential Soil Microbial EcologyRating: 5 out of 5 stars5/5 (1)
- How People Make Their Own EnvironmentDocument13 pagesHow People Make Their Own EnvironmentAlfin HasanNo ratings yet
- 2021 Bookmatter Plant-AnimalInteractionsDocument13 pages2021 Bookmatter Plant-AnimalInteractionsAna Clara P. TeixeiraNo ratings yet
- Gross 2005Document11 pagesGross 2005pomajoluNo ratings yet
- Scarr McCartneyDocument13 pagesScarr McCartneyRocío Donaire CortésNo ratings yet
- Approaches, and A Proposal For A Future Research StrategyDocument15 pagesApproaches, and A Proposal For A Future Research StrategyJoão Marcelo Brazão ProtázioNo ratings yet
- Liu2020 Phenotypic Plasticity Contributes To Maize Adaptation and HeterosisDocument14 pagesLiu2020 Phenotypic Plasticity Contributes To Maize Adaptation and HeterosisDuc Nguyen TrungNo ratings yet
- EcotoxicologiaDocument1 pageEcotoxicologiaCatalina PetrelNo ratings yet
- Manjit S. Kang, Hugh G. Gauch-Genotype - by - Environment Interaction PDFDocument409 pagesManjit S. Kang, Hugh G. Gauch-Genotype - by - Environment Interaction PDFLenio Urzeda100% (3)
- What Determines Perceived Income JusticeDocument33 pagesWhat Determines Perceived Income JusticeBoagiu RăzvanNo ratings yet
- 18 - Murren (2015) Limits and Costs of Phenotype and PlasticityDocument9 pages18 - Murren (2015) Limits and Costs of Phenotype and PlasticityClaivert FelixNo ratings yet
- Horne2019 Article ASynthesisOfMajorEnvironmentalDocument11 pagesHorne2019 Article ASynthesisOfMajorEnvironmentalMellya RizkiNo ratings yet
- Corral-Verdugo2002 MODELO COMPETENCIAS AMBIENTALESDocument19 pagesCorral-Verdugo2002 MODELO COMPETENCIAS AMBIENTALESDanielaNo ratings yet
- Laboratory Exercises-EcologyDocument12 pagesLaboratory Exercises-EcologyAntonia Mikaela BagasNo ratings yet
- KamelOyarzunGrosberg 2010Document12 pagesKamelOyarzunGrosberg 2010fygv1990No ratings yet
- Corruption of Journal Impact Factors: Anurag A. AgrawalDocument2 pagesCorruption of Journal Impact Factors: Anurag A. AgrawalArmansyah maulana harahapNo ratings yet
- Orrock Et Al JB 2015Document9 pagesOrrock Et Al JB 2015ronaldoNo ratings yet
- Differential Effects of Developmental Thermal PlasDocument12 pagesDifferential Effects of Developmental Thermal PlasAlan David Ruiz GarcíaNo ratings yet
- Knafonoam 2017Document26 pagesKnafonoam 2017Maria-Magdalena AbracelNo ratings yet
- BetweenDocument18 pagesBetweenj.k.steinbergerNo ratings yet
- Frontiers Ecology: in and TheDocument9 pagesFrontiers Ecology: in and TheJhonatan GutierrezNo ratings yet
- Call Away Etal 2003 EcologyDocument15 pagesCall Away Etal 2003 EcologyMickaël Houessou SononNo ratings yet
- Parental Effects and Climate Change Will Avian IncubationDocument13 pagesParental Effects and Climate Change Will Avian IncubationAyu Liani VeronikaNo ratings yet
- 1 s2.0 S0003996908001842 MainDocument7 pages1 s2.0 S0003996908001842 MainAdrienn MatheNo ratings yet
- Shared Environment 2021Document9 pagesShared Environment 2021valericsamNo ratings yet
- 1 s2.0 S1439179121001857 MainDocument12 pages1 s2.0 S1439179121001857 Mainjafar-fpmNo ratings yet
- Hesse Et Al-2015-Journal of Evolutionary BiologyDocument9 pagesHesse Et Al-2015-Journal of Evolutionary BiologySam ChenNo ratings yet
- Biodiversity and Ecosystem Function Science 2012 Jan 335-6065 - 174-5Document3 pagesBiodiversity and Ecosystem Function Science 2012 Jan 335-6065 - 174-5Unram OfficialNo ratings yet
- Does Ecology Need A Technical Revolution - Honours Course Extinction - Sebastiaan de Jager 11301430Document5 pagesDoes Ecology Need A Technical Revolution - Honours Course Extinction - Sebastiaan de Jager 11301430api-408361250No ratings yet
- Environmental Research: A B F C B G A B D A A A e ADocument12 pagesEnvironmental Research: A B F C B G A B D A A A e ABen AllenNo ratings yet
- M. Aldana (2020)Document8 pagesM. Aldana (2020)steeven.j.alejandroNo ratings yet
- Thesis Statement On GM FoodsDocument4 pagesThesis Statement On GM Foodsaflpaftaofqtoa100% (2)
- Weller Et Al 2023 New Insight Into Drivers of Mammalian Litter Size From Individual Level TraitsDocument14 pagesWeller Et Al 2023 New Insight Into Drivers of Mammalian Litter Size From Individual Level TraitsNorberto MuzzachiodiNo ratings yet
- Final Exam Environmental BiologyDocument10 pagesFinal Exam Environmental BiologyMagy GuzmanNo ratings yet
- Agronomy Journal - 2019 - Olivoto - Mean Performance and Stability in Multi Environment Trials II Selection Based OnDocument9 pagesAgronomy Journal - 2019 - Olivoto - Mean Performance and Stability in Multi Environment Trials II Selection Based OnGisele HolandaNo ratings yet
- Eco Metaanaliza Human Connection To Nature and Proenvironmental Behavior Whitburn, Linklater, AbrahamseDocument14 pagesEco Metaanaliza Human Connection To Nature and Proenvironmental Behavior Whitburn, Linklater, AbrahamseKarolina JackiewiczNo ratings yet
- Life Span Human Development 9th Edition Sigelman Solutions ManualDocument19 pagesLife Span Human Development 9th Edition Sigelman Solutions ManualJessicaPorteraiydm100% (16)
- Costanza 2007 - Biodiversity and Ecosystem ServicesDocument14 pagesCostanza 2007 - Biodiversity and Ecosystem ServicesGuilherme PradoNo ratings yet
- Plant Life History and Above-Belowground Interactions: Missing LinksDocument11 pagesPlant Life History and Above-Belowground Interactions: Missing LinksrachiiidaNo ratings yet
- Hurst Dittmar Bond Kasser 2013Document13 pagesHurst Dittmar Bond Kasser 2013Belhadj TarekNo ratings yet
- Zekeng Et Al 2023Document13 pagesZekeng Et Al 2023Citoyen x14No ratings yet
- Ddi 12970Document12 pagesDdi 12970Vellin LusianaNo ratings yet
- Costa Etal 2018 GEBDocument14 pagesCosta Etal 2018 GEBpcorgoNo ratings yet
- Melo Etal 2017 Testing Habitat Amount Hypothesis Biol Conserv PDFDocument12 pagesMelo Etal 2017 Testing Habitat Amount Hypothesis Biol Conserv PDFgbitencourt.bioNo ratings yet
- Bio 4Document45 pagesBio 4vmanimaranNo ratings yet
- Ernst + Banks (20 Humans Integrate Visual and Haptic Information in A Statistically Optimal FashionDocument5 pagesErnst + Banks (20 Humans Integrate Visual and Haptic Information in A Statistically Optimal FashionCosnita Emilia-FeliciaNo ratings yet
- Pecquerie 2009Document13 pagesPecquerie 2009Fadhli LatuconsinaNo ratings yet
- Grime 1998 J EcolDocument10 pagesGrime 1998 J EcolAlex GrünlichNo ratings yet
- Dwnload Full Life Span Human Development 9th Edition Sigelman Solutions Manual PDFDocument35 pagesDwnload Full Life Span Human Development 9th Edition Sigelman Solutions Manual PDFbeliefknackq3zcb100% (9)
- Behavioral Plasticity. An Interaction Between Evolution and ExperienceDocument13 pagesBehavioral Plasticity. An Interaction Between Evolution and ExperiencevmsolartecNo ratings yet
- Economic Growth, Biodiversity Loss and Conservation Effort: Simon Dietz, W. Neil AdgerDocument13 pagesEconomic Growth, Biodiversity Loss and Conservation Effort: Simon Dietz, W. Neil AdgerQamar AliNo ratings yet
- The Use of Telomerized Cells For Tissue Engineering: AnalysisDocument2 pagesThe Use of Telomerized Cells For Tissue Engineering: AnalysisDaniela Mădălina GhețuNo ratings yet
- Giling - 2018 - GCB - A Niche For Ecosystem Multifunctionality in Global Change ResearchDocument12 pagesGiling - 2018 - GCB - A Niche For Ecosystem Multifunctionality in Global Change ResearchJenickson CostaNo ratings yet
- Tmpa851 TMPDocument9 pagesTmpa851 TMPFrontiersNo ratings yet
- AAB-Uphoffetal 2009Document27 pagesAAB-Uphoffetal 2009Usama TareenNo ratings yet
- CH 1.1 - 1.5 The Environment and You Caterina AbelDocument8 pagesCH 1.1 - 1.5 The Environment and You Caterina AbelCaterina AbelNo ratings yet
- Grass Research PaperDocument4 pagesGrass Research Paperajqkrxplg100% (1)
- Annotated BibliographyDocument3 pagesAnnotated Bibliographyapi-486758542No ratings yet
- A Scoping Review of The Impact On Children of The Built Environment Design Characteristics of Healing SpacesDocument18 pagesA Scoping Review of The Impact On Children of The Built Environment Design Characteristics of Healing SpacesRevielyn RoseNo ratings yet
- Taylor Hughes ThesisDocument8 pagesTaylor Hughes Thesislindatorrespaterson100% (2)
- Germination and Post Germination Growth To Carrot Seedling Emergence: Predictive Threshold Models and Sources of Variation Between Sowing OccasionsDocument12 pagesGermination and Post Germination Growth To Carrot Seedling Emergence: Predictive Threshold Models and Sources of Variation Between Sowing OccasionsAndre LanzerNo ratings yet
- Predictive Mapping of Alpine Grasslands in Switzerland - Species Versus Community ApproachDocument14 pagesPredictive Mapping of Alpine Grasslands in Switzerland - Species Versus Community ApproachAndre LanzerNo ratings yet
- A Simulation Model For The Transient Effects of Climate Change On Forest LandscapesDocument20 pagesA Simulation Model For The Transient Effects of Climate Change On Forest LandscapesAndre LanzerNo ratings yet
- Improving The Behaviour of Forest Gap Models Along Drought GradientsDocument18 pagesImproving The Behaviour of Forest Gap Models Along Drought GradientsAndre LanzerNo ratings yet
- Monitoring Vegetation Systems in The Great Plains With ERTSDocument25 pagesMonitoring Vegetation Systems in The Great Plains With ERTSAndre LanzerNo ratings yet
- Book Citations: Influence of Epidemiologic Thought in The Academic CommunityDocument7 pagesBook Citations: Influence of Epidemiologic Thought in The Academic CommunityAndre LanzerNo ratings yet
- What Properties Might Statistical Inferences Reasonably Be Expected To Have ? Crisis and Resolution in Statistical InferenceDocument11 pagesWhat Properties Might Statistical Inferences Reasonably Be Expected To Have ? Crisis and Resolution in Statistical InferenceAndre LanzerNo ratings yet
- Advances in Quantitative Methods in EpidemiologyDocument8 pagesAdvances in Quantitative Methods in EpidemiologyAndre LanzerNo ratings yet
- The Effects of Species Range Sizes On The Accuracy of Distribution Models: Ecological Phenomenon or Statistical Artefact ?Document13 pagesThe Effects of Species Range Sizes On The Accuracy of Distribution Models: Ecological Phenomenon or Statistical Artefact ?Andre LanzerNo ratings yet
- Observational Studies and Bias in EpidemiologyDocument36 pagesObservational Studies and Bias in EpidemiologyAndre LanzerNo ratings yet
- Observational Studies: Why Are They So ImportantDocument2 pagesObservational Studies: Why Are They So ImportantAndre LanzerNo ratings yet
- A Review of Models of Landscape ChangeDocument23 pagesA Review of Models of Landscape ChangeAndre LanzerNo ratings yet
- The Optimal Balance Between Size and Number of OffspringDocument9 pagesThe Optimal Balance Between Size and Number of OffspringAndre LanzerNo ratings yet
- That Confounded P-ValueDocument2 pagesThat Confounded P-ValueAndre LanzerNo ratings yet
- Selection Bias Using Observed Data Augmented With Imputed Record-Level Probabilities.Document18 pagesSelection Bias Using Observed Data Augmented With Imputed Record-Level Probabilities.Andre LanzerNo ratings yet
- Evolution of Epidemiologic Methods and Concepts in Selected Textbooks of The 20th CenturyDocument9 pagesEvolution of Epidemiologic Methods and Concepts in Selected Textbooks of The 20th CenturyAndre LanzerNo ratings yet
- The Evidence-Based Health Care Debate - 2006. Where Are We Now ?Document9 pagesThe Evidence-Based Health Care Debate - 2006. Where Are We Now ?Andre LanzerNo ratings yet
- Evidence-Based Health Policy: A Preliminary Systematic ReviewDocument5 pagesEvidence-Based Health Policy: A Preliminary Systematic ReviewAndre LanzerNo ratings yet
- Methods in Epidemiology and Public Health: Does Practice Match Theory ?Document7 pagesMethods in Epidemiology and Public Health: Does Practice Match Theory ?Andre LanzerNo ratings yet
- An Introduction To The Use of Epidemiologic Research Methods in Dairy ScienceDocument8 pagesAn Introduction To The Use of Epidemiologic Research Methods in Dairy ScienceAndre LanzerNo ratings yet
- Bryant, P. Brown, R. and Abarbanel, H.D.I. Lyapunov Exponents From Observed Time Series. Physical Review Letters 65 (13) : 1523-1526. 1990.Document4 pagesBryant, P. Brown, R. and Abarbanel, H.D.I. Lyapunov Exponents From Observed Time Series. Physical Review Letters 65 (13) : 1523-1526. 1990.Andre LanzerNo ratings yet
- Rouse (1974) - MONITORING VEGETATION SYSTEMS IN THE GREAT PLAINS WITH ERTSDocument9 pagesRouse (1974) - MONITORING VEGETATION SYSTEMS IN THE GREAT PLAINS WITH ERTSAndre LanzerNo ratings yet
- Scale Effects in Landscape StudiesDocument20 pagesScale Effects in Landscape StudiesAndre LanzerNo ratings yet
- Evidence-Based Public Health: A Fundamental Concept For Public Health PracticeDocument30 pagesEvidence-Based Public Health: A Fundamental Concept For Public Health PracticeAndre LanzerNo ratings yet
- Modeling and Variable Selection in Epidemiologic AnalysisDocument10 pagesModeling and Variable Selection in Epidemiologic AnalysisAndre LanzerNo ratings yet
- Simple Mathematical Models With Very Complicated DynamicsDocument10 pagesSimple Mathematical Models With Very Complicated DynamicsAndre LanzerNo ratings yet
- The Coefficient of Determinaton Exposed - HahnDocument4 pagesThe Coefficient of Determinaton Exposed - HahnAndre LanzerNo ratings yet
- Fractals and EcologyDocument18 pagesFractals and EcologyAndre LanzerNo ratings yet
- Markov Chain: A Realistic Model For Temporal Coenosere ?Document24 pagesMarkov Chain: A Realistic Model For Temporal Coenosere ?Andre LanzerNo ratings yet
- C57.12.29 Pad-Mounted Equipment-Enclosure Integrity For Coastal EnvironmentsDocument26 pagesC57.12.29 Pad-Mounted Equipment-Enclosure Integrity For Coastal EnvironmentsSureshKumarNo ratings yet
- Employment Under Dubai Electricity & Water AuthorityDocument6 pagesEmployment Under Dubai Electricity & Water AuthorityMominur Rahman ShohagNo ratings yet
- Waste Heat BoilerDocument7 pagesWaste Heat Boilerabdul karimNo ratings yet
- Universal Human Values NotesDocument13 pagesUniversal Human Values NotesS RamNo ratings yet
- THDP-3 Worm Gear Manual Interchangeable Single Punch Tablet PressDocument8 pagesTHDP-3 Worm Gear Manual Interchangeable Single Punch Tablet Pressdibimrane13No ratings yet
- Jastram Rudder Feedback UnitDocument21 pagesJastram Rudder Feedback UnitGary Gouveia100% (3)
- Embargoed: Embargoed Until April 24, 2019 at 12:01 A.M. (Eastern Time)Document167 pagesEmbargoed: Embargoed Until April 24, 2019 at 12:01 A.M. (Eastern Time)Las Vegas Review-JournalNo ratings yet
- Dossat PrinciplesOfRefrigerationDocument554 pagesDossat PrinciplesOfRefrigerationHernan CobaNo ratings yet
- Long PassagesDocument12 pagesLong PassagesAml AmlNo ratings yet
- KIT REQUEST FORM - GradDocument2 pagesKIT REQUEST FORM - Graddamie aadamsNo ratings yet
- Bibliografi 3Document3 pagesBibliografi 3Praba RauNo ratings yet
- Seventh Pay Commission ArrearsDocument11 pagesSeventh Pay Commission Arrearssantosh bharathyNo ratings yet
- Faculty of Civil Engineering and Planning Civil Engineering Department Petra Christian UniversityDocument59 pagesFaculty of Civil Engineering and Planning Civil Engineering Department Petra Christian Universitykenny lieNo ratings yet
- Perkins - General Ti BulletinDocument65 pagesPerkins - General Ti BulletinUTEL CARTERNo ratings yet
- The Roosh ProgramDocument2 pagesThe Roosh ProgramVal KerryNo ratings yet
- ECON 304 Course ContentDocument2 pagesECON 304 Course ContentAanand JhaNo ratings yet
- SAGC Compliance Awareness-Grid UsersDocument66 pagesSAGC Compliance Awareness-Grid Userskamal_khan85No ratings yet
- Loading and Hauling Dump Truck: DumperDocument2 pagesLoading and Hauling Dump Truck: DumperRJNo ratings yet
- 01 AirCircuitBreakers LV10 102021 EN 202111301146249676Document136 pages01 AirCircuitBreakers LV10 102021 EN 202111301146249676bkNo ratings yet
- Ens Air To Water Operation ManualDocument8 pagesEns Air To Water Operation ManualcomborNo ratings yet
- Green ICT: A Study of Awareness, Attitude and Adoption Among IT/Computer Engineering Students of LDRP-ITR, GandhinagarDocument13 pagesGreen ICT: A Study of Awareness, Attitude and Adoption Among IT/Computer Engineering Students of LDRP-ITR, GandhinagarAHMAD ARESYADNo ratings yet
- Vernacular in Andhra PradeshDocument1 pageVernacular in Andhra PradeshNandyala Rajarajeswari DeviNo ratings yet
- Quotation Request Form: Customer DetailsDocument1 pageQuotation Request Form: Customer DetailsAmanda RezendeNo ratings yet
- Gaara Hiden Series Light NovelDocument127 pagesGaara Hiden Series Light NovelartfardadNo ratings yet
- Switch RotatoriosDocument12 pagesSwitch RotatoriosRoberto AndrésNo ratings yet
- G120D Getting Started 0418 en-USDocument94 pagesG120D Getting Started 0418 en-USHamadi Ben SassiNo ratings yet
- Leap Motion PDFDocument18 pagesLeap Motion PDFAnkiTwilightedNo ratings yet
- Engineering Properties (Al O) : 94% Aluminum Oxide Mechanical Units of Measure SI/Metric (Imperial)Document7 pagesEngineering Properties (Al O) : 94% Aluminum Oxide Mechanical Units of Measure SI/Metric (Imperial)Hendy SetiawanNo ratings yet
- Transient Thermal Conduction ExampleDocument14 pagesTransient Thermal Conduction ExampleDevendra BangarNo ratings yet
- CS221 - Artificial Intelligence - Search - 4 Dynamic ProgrammingDocument23 pagesCS221 - Artificial Intelligence - Search - 4 Dynamic ProgrammingArdiansyah Mochamad NugrahaNo ratings yet