You might also like
- Arthropod Structure & Development: Fabian B Aumler, Stanislav N. Gorb, Sebastian BüsseDocument12 pagesArthropod Structure & Development: Fabian B Aumler, Stanislav N. Gorb, Sebastian BüsseSuhar TatiNo ratings yet
- VanderLinden Et Al 2019 Head Turning MorphologiesDocument12 pagesVanderLinden Et Al 2019 Head Turning MorphologiesKo KaNo ratings yet
- Abdala Et Al 2015 J Morph 276Document21 pagesAbdala Et Al 2015 J Morph 276mioclaenusNo ratings yet
- Mui Et Al 2012Document13 pagesMui Et Al 2012api-263358650No ratings yet
- Rat AnatomiaDocument42 pagesRat AnatomiaKarina Fischer SigelNo ratings yet
- Journal of AnatomyDocument14 pagesJournal of AnatomyAlejo RuilovaNo ratings yet
- VOLUME MUSCULAR - Upper Limb Muscle Volumes in Adult Subjects - 2007 PDFDocument8 pagesVOLUME MUSCULAR - Upper Limb Muscle Volumes in Adult Subjects - 2007 PDFFernando De Oliveira SolimanNo ratings yet
- 3D imaging reveals anatomy of male genital organs in red foxDocument10 pages3D imaging reveals anatomy of male genital organs in red foxAva DávidNo ratings yet
- 2006 - Sensory Control of Extraocular MusclesDocument13 pages2006 - Sensory Control of Extraocular MusclesFede NahasNo ratings yet
- Are Endocasts Good Proxies For Brain Size and Shape inDocument15 pagesAre Endocasts Good Proxies For Brain Size and Shape inlola.carvallhoNo ratings yet
- ArtigoDocument10 pagesArtigocaroline.sottoNo ratings yet
- The Anatomical Record - 2009 - Holliday - New Insights Into Dinosaur Jaw Muscle AnatomyDocument20 pagesThe Anatomical Record - 2009 - Holliday - New Insights Into Dinosaur Jaw Muscle AnatomyKidus TarekegnNo ratings yet
- Pucciarelli 2000Document12 pagesPucciarelli 2000shikarineha5591No ratings yet
- Tingle Et Al, 2023Document20 pagesTingle Et Al, 2023Livia PereiraNo ratings yet
- Comparative AnatomyDocument38 pagesComparative AnatomyKimm Charmaine RodriguezNo ratings yet
- Form, Function, Food and Feeding in Stellate Echinoderms : ZoosymposiaDocument10 pagesForm, Function, Food and Feeding in Stellate Echinoderms : ZoosymposiaSixto Gutiérrez SaavedraNo ratings yet
- Artículo Original/ Original ArticleDocument9 pagesArtículo Original/ Original ArticleCami ChavezNo ratings yet
- GIFFORD-GONZALEZ. 1989. Ethnographic Analogues For Interpreting Modified Bones Some Cases From East Africa PDFDocument68 pagesGIFFORD-GONZALEZ. 1989. Ethnographic Analogues For Interpreting Modified Bones Some Cases From East Africa PDFFrancisco Caro OlivaresNo ratings yet
- Mastication Muscles in Hoary Fox (Lycalopex Vetulus - LUND, 1842) : Descriptive and Comparative StudyDocument9 pagesMastication Muscles in Hoary Fox (Lycalopex Vetulus - LUND, 1842) : Descriptive and Comparative StudyIJAERS JOURNALNo ratings yet
- Journal of Human Evolution: Gabrielle A. Russo, Liza J. ShapiroDocument13 pagesJournal of Human Evolution: Gabrielle A. Russo, Liza J. ShapiroYsNo ratings yet
- The Articular Muscle of The Knee: Morphology and DispositionDocument4 pagesThe Articular Muscle of The Knee: Morphology and DispositionFelipe SánchezNo ratings yet
- Organogenesis of The Musculoskeletal System in Horse Embryos and Early FetusesDocument8 pagesOrganogenesis of The Musculoskeletal System in Horse Embryos and Early FetusesMackarenna NarvaezNo ratings yet
- The Use of Otolith Morphometrics in Determining The Size and Species Identification of Eight Mullets (Mugiliformes: Mugilidae) From MalaysiaDocument9 pagesThe Use of Otolith Morphometrics in Determining The Size and Species Identification of Eight Mullets (Mugiliformes: Mugilidae) From MalaysiaUkki Aishi NyoNo ratings yet
- Abdala Et Al 2008 J Anat 213Document14 pagesAbdala Et Al 2008 J Anat 213mioclaenusNo ratings yet
- De Groote and Humphey 2011 Masa Corp Metatarso1Document8 pagesDe Groote and Humphey 2011 Masa Corp Metatarso1Keila MenaNo ratings yet
- 2017 Gsiorowski Et AlDocument29 pages2017 Gsiorowski Et AlDayana FlorezNo ratings yet
- Relationships Between the Size and Spatial Morphology of human Masseter and medial Pterygoid muscles- the Craniofacial skelton and jaw mechanics.pdf مهم جداDocument17 pagesRelationships Between the Size and Spatial Morphology of human Masseter and medial Pterygoid muscles- the Craniofacial skelton and jaw mechanics.pdf مهم جداد.عاصم صويلحNo ratings yet
- Elife 43828Document19 pagesElife 43828jaypasco79No ratings yet
- Origens e Ramificações Do Plexo Braquial No Cachorro-Do-MatoDocument13 pagesOrigens e Ramificações Do Plexo Braquial No Cachorro-Do-MatoAnna Luiza AraújoNo ratings yet
- Cranial Muscles of South American GeckosDocument12 pagesCranial Muscles of South American GeckosmioclaenusNo ratings yet
- The Anatomical Record - 2022 - Richards - Endocranial Anatomy of The Guercy 1 Early Neanderthal From Baume Moula GuercyDocument30 pagesThe Anatomical Record - 2022 - Richards - Endocranial Anatomy of The Guercy 1 Early Neanderthal From Baume Moula Guercylola.carvallhoNo ratings yet
- Quantitative and Descriptive Analysis of The Meniscotibial Ligament in Human CorpsesDocument3 pagesQuantitative and Descriptive Analysis of The Meniscotibial Ligament in Human CorpsesSaifuddin HaswareNo ratings yet
- Journal of Anatomy - 2019 - Echeverr ADocument13 pagesJournal of Anatomy - 2019 - Echeverr Apjames199705No ratings yet
- Histomorphometric Evaluation of The Effects of Ovariectomy On Bone Turnover in Rat Caudal VertebraeDocument7 pagesHistomorphometric Evaluation of The Effects of Ovariectomy On Bone Turnover in Rat Caudal VertebraeSara HernandezNo ratings yet
- Morphological and Topographical Anatomy of Nutrient Foramen in The Lower Limb Long BonesDocument8 pagesMorphological and Topographical Anatomy of Nutrient Foramen in The Lower Limb Long Bonesmiskica miskicaNo ratings yet
- The Caudal Skeleton of Extant and Fossil Cyprinodontiform Fishes A Comparative MorphologyDocument20 pagesThe Caudal Skeleton of Extant and Fossil Cyprinodontiform Fishes A Comparative MorphologyAquaponics OaxacaNo ratings yet
- 2012 Article 148Document9 pages2012 Article 148João GasparNo ratings yet
- Tendon Del Subescapular - Sistema de Clasificación - AnatomíaDocument7 pagesTendon Del Subescapular - Sistema de Clasificación - AnatomíajesusglezctNo ratings yet
- Shape and Volume of Craniofacial Cavities in Intentional Skull DeformationsDocument10 pagesShape and Volume of Craniofacial Cavities in Intentional Skull DeformationsGastón SalasNo ratings yet
- Geospiza Bite ForceDocument13 pagesGeospiza Bite ForceHappyMark93No ratings yet
- 2014 The Anatomy of The Hip Abductors MusclesDocument13 pages2014 The Anatomy of The Hip Abductors Musclesmari netaNo ratings yet
- 2012 Oviraptorosaur Tail Forms and FunctionsDocument47 pages2012 Oviraptorosaur Tail Forms and FunctionsJorge BarrientosNo ratings yet
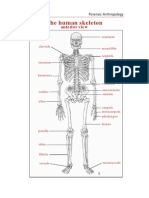
- The Human Skeleton: Anterior ViewDocument0 pagesThe Human Skeleton: Anterior VieweduardoarcticNo ratings yet
- 2012 RPVBDocument5 pages2012 RPVBRafael MaiorNo ratings yet
- Abdominal Skeleton Variation in Tropidurid LizardsDocument11 pagesAbdominal Skeleton Variation in Tropidurid Lizardsparabin.3528101No ratings yet
- Paxinos and Franklin's the Mouse Brain in Stereotaxic Coordinates, Compact: The Coronal Plates and DiagramsFrom EverandPaxinos and Franklin's the Mouse Brain in Stereotaxic Coordinates, Compact: The Coronal Plates and DiagramsNo ratings yet
- Anatomia Metatarso FalangeDocument9 pagesAnatomia Metatarso FalangeAmairany García PeñaNo ratings yet
- Zoology MJ-2 MJ-3Document6 pagesZoology MJ-2 MJ-3vishalbisai07No ratings yet
- Ratification Page: A Citra Pratiwi, S.PD, M. Ed Muhammad Nur Arsyad. S.PDDocument13 pagesRatification Page: A Citra Pratiwi, S.PD, M. Ed Muhammad Nur Arsyad. S.PDbertha tandiNo ratings yet
- Evolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesDocument11 pagesEvolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesKo KaNo ratings yet
- Adductor Magnus An EMG Investigation Into Proximal and Disctal Portions and Direction Specific ActionDocument9 pagesAdductor Magnus An EMG Investigation Into Proximal and Disctal Portions and Direction Specific ActionElaine CspNo ratings yet
- HHS Public Access: Three-Dimensional Stiffness of The Carpal ArchDocument14 pagesHHS Public Access: Three-Dimensional Stiffness of The Carpal ArcherlysaNo ratings yet
- This Content Downloaded From 132.247.111.165 On Tue, 20 Sep 2022 20:09:23 UTCDocument15 pagesThis Content Downloaded From 132.247.111.165 On Tue, 20 Sep 2022 20:09:23 UTCKarina SjNo ratings yet
- The Anatomy of The Articularis Genus Muscle and ItDocument7 pagesThe Anatomy of The Articularis Genus Muscle and ItTodirenche LarisaNo ratings yet
- Articularis Genus: An Anatomic and MRI Study in CadaversDocument9 pagesArticularis Genus: An Anatomic and MRI Study in CadaversFelipe SánchezNo ratings yet
- Anatomical Study On The Anterolateral Ligament of The Knee: Original ArticleDocument6 pagesAnatomical Study On The Anterolateral Ligament of The Knee: Original ArticleAngel ArturoNo ratings yet
- Skull and Fin Bone Development in Yellowtail FishDocument13 pagesSkull and Fin Bone Development in Yellowtail FishKATALINA IGNACIA AZÓCARNo ratings yet
- New Gecko Species from SulawesiDocument11 pagesNew Gecko Species from Sulawesiheru2910No ratings yet
- Biomechanics Pronation SupinationDocument10 pagesBiomechanics Pronation SupinationProfesseur Christian DumontierNo ratings yet
- The Effect of Hyaluronidase On Depth of Necrosis in Hyaluronic Acid Filling-Related Skin ComplicationsDocument8 pagesThe Effect of Hyaluronidase On Depth of Necrosis in Hyaluronic Acid Filling-Related Skin ComplicationsKerlida SantosNo ratings yet
- Complications Secondary To Nonsurgical Rhinoplasty A Systematic Review and Meta-Analysis - de Victor, 2021Document6 pagesComplications Secondary To Nonsurgical Rhinoplasty A Systematic Review and Meta-Analysis - de Victor, 2021Rafael Autran Cavalcante AraújoNo ratings yet
- Clinical Outcomes of Impending Nasal Skin Necrosis Related To Nose and Nasolabial Fold Augmentation With Hyaluronic Acid FillersDocument8 pagesClinical Outcomes of Impending Nasal Skin Necrosis Related To Nose and Nasolabial Fold Augmentation With Hyaluronic Acid FillersKerlida SantosNo ratings yet
- Treatment of Skin Soft Tissue Embolism After Hyaluronic Acid Injection For Injection Rhinoplasty in Asian PatientsDocument8 pagesTreatment of Skin Soft Tissue Embolism After Hyaluronic Acid Injection For Injection Rhinoplasty in Asian PatientsKerlida SantosNo ratings yet
- Cerebral Embolism As A Result of Facial Filler Injections - A Literature ReviewDocument14 pagesCerebral Embolism As A Result of Facial Filler Injections - A Literature ReviewKerlida SantosNo ratings yet
- Color Doppler Flow Imaging of Retrobulbar Ocular Blood Flow Changes in Retinal Artery Occlusions Caused by Cosmetic Facial Filler InjectionsDocument5 pagesColor Doppler Flow Imaging of Retrobulbar Ocular Blood Flow Changes in Retinal Artery Occlusions Caused by Cosmetic Facial Filler InjectionsKerlida SantosNo ratings yet
- Complete Recovery of Filler-Induced Visual Loss Following Subcutaneous Hyaluronidase InjectionDocument5 pagesComplete Recovery of Filler-Induced Visual Loss Following Subcutaneous Hyaluronidase InjectionKerlida SantosNo ratings yet
- OSCE Checklist - Hernia Examination-1Document1 pageOSCE Checklist - Hernia Examination-1singhurvashi365No ratings yet
- OSCE Checklist Oral Cavity Examination 1Document2 pagesOSCE Checklist Oral Cavity Examination 1Kerlida SantosNo ratings yet
- Mental Capacity AssessmentDocument1 pageMental Capacity AssessmentKerlida SantosNo ratings yet
- OSCE Checklist - Hernia Examination-1Document1 pageOSCE Checklist - Hernia Examination-1singhurvashi365No ratings yet
- OSCE Checklist Oral Cavity Examination 1Document2 pagesOSCE Checklist Oral Cavity Examination 1Kerlida SantosNo ratings yet
- Cannula Versus Needle in Medical Rhinoplasty - The Nose KnowsDocument24 pagesCannula Versus Needle in Medical Rhinoplasty - The Nose KnowsKerlida SantosNo ratings yet
- Avoiding and Treating Blindness From Fillers: A Review of The World LiteratureDocument21 pagesAvoiding and Treating Blindness From Fillers: A Review of The World LiteratureSam CruzNo ratings yet
- Panophthalmoplegia and Vision Loss After Cosmetic Nasal Dorsum InjectionDocument3 pagesPanophthalmoplegia and Vision Loss After Cosmetic Nasal Dorsum InjectionKerlida SantosNo ratings yet
- OSCE Checklist Cerebellar ExaminationDocument2 pagesOSCE Checklist Cerebellar ExaminationKerlida SantosNo ratings yet
- OSCE Checklist Otoscopy and Hearing AssessmentDocument2 pagesOSCE Checklist Otoscopy and Hearing AssessmentKerlida SantosNo ratings yet
- OSCE Checklist Breast ExaminationDocument1 pageOSCE Checklist Breast ExaminationKerlida Santos100% (1)
- Patient Satisfaction After Non-Surgical Rhinoplasty Using Hyaluronic Acid - A Literature ReviewDocument6 pagesPatient Satisfaction After Non-Surgical Rhinoplasty Using Hyaluronic Acid - A Literature ReviewKerlida SantosNo ratings yet
- OSCE Checklist Varicose Vein ExaminationDocument2 pagesOSCE Checklist Varicose Vein ExaminationKerlida SantosNo ratings yet
- A Safer Non-surgical Filler Augmentation Rhinoplasty TechniqueDocument6 pagesA Safer Non-surgical Filler Augmentation Rhinoplasty TechniqueАндрей ПетровNo ratings yet
- Effects of N-Acetyl-cysteine On The Survival and Regeneration ofDocument9 pagesEffects of N-Acetyl-cysteine On The Survival and Regeneration ofKerlida SantosNo ratings yet
- The ENT History and ExaminationDocument11 pagesThe ENT History and ExaminationSuresh MettaNo ratings yet
- Irfan Mir Notes (Mubasher Abbas - bnp1000)Document16 pagesIrfan Mir Notes (Mubasher Abbas - bnp1000)usmle1No ratings yet
- Irfan Mir Notes (Mubasher Abbas - bnp1000)Document16 pagesIrfan Mir Notes (Mubasher Abbas - bnp1000)usmle1No ratings yet
- Ancient Genomic Time Transect From The Central Asian Steppe Unravels The History of The ScythiansDocument15 pagesAncient Genomic Time Transect From The Central Asian Steppe Unravels The History of The ScythiansErwin SchrödringerNo ratings yet
- Notes of Chapter 1 Biology Class 12 (Reproduction in Organisms)Document4 pagesNotes of Chapter 1 Biology Class 12 (Reproduction in Organisms)Abhay GautamNo ratings yet
- Keanekaragaman Dan Biomassa Ikan Karang Serta Keterkaitannya Dengan Tutupan Karang Hidup Di Perairan Manggis, Kabupaten Karangasem, BaliDocument13 pagesKeanekaragaman Dan Biomassa Ikan Karang Serta Keterkaitannya Dengan Tutupan Karang Hidup Di Perairan Manggis, Kabupaten Karangasem, BalikhanphitaNo ratings yet
- The Mothers Matriarchal Theory Social Origins Robert Briffault 1931Document334 pagesThe Mothers Matriarchal Theory Social Origins Robert Briffault 1931Rene GonzalezNo ratings yet
- Charles Darwin: On The Origin of SpeciesDocument3 pagesCharles Darwin: On The Origin of SpeciesJosip MihicNo ratings yet
- Tugas Bahasa Inggris-1Document8 pagesTugas Bahasa Inggris-1Nur KomariyahNo ratings yet
- مطوية PDFDocument2 pagesمطوية PDFAbdou VerrattiNo ratings yet
- Williams Syndrome Research PaperDocument7 pagesWilliams Syndrome Research Paperefh4m77n100% (1)
- Hematology AutomationDocument117 pagesHematology AutomationRico EdureseNo ratings yet
- Biochemistry TA Notes Part 1Document49 pagesBiochemistry TA Notes Part 1lovelyc95No ratings yet
- Evs Notes - Unit IDocument7 pagesEvs Notes - Unit Iwolfy shadow8No ratings yet
- Stedman's AnatomyDocument31 pagesStedman's AnatomyHein Aung ZawNo ratings yet
- Genetics Notes: Independent Assortment No Effect Father of GeneticsDocument22 pagesGenetics Notes: Independent Assortment No Effect Father of GeneticsTevfik DoğanNo ratings yet
- What Affects The Biocompatibility of PolymersDocument25 pagesWhat Affects The Biocompatibility of PolymersAndreaNo ratings yet
- Johnson 2003Document47 pagesJohnson 2003Sixto Gutiérrez SaavedraNo ratings yet
- Gambar Klasifikasi Kelinci Lop: KingdomDocument4 pagesGambar Klasifikasi Kelinci Lop: Kingdomkiana100% (1)
- 1 PBDocument10 pages1 PBDwi Rama NugrahaNo ratings yet
- Diseases of Mites and TicksDocument344 pagesDiseases of Mites and TicksMaria Eduarda Chiaria FurquimNo ratings yet
- Buthidae Family Overview: Taxonomy and Genera of the Largest Scorpion FamilyDocument54 pagesButhidae Family Overview: Taxonomy and Genera of the Largest Scorpion FamilyAlejandro Tampoa0% (1)
- Post Test Structure and Written Expression ToeflDocument7 pagesPost Test Structure and Written Expression ToeflfaniNo ratings yet
- Don Bosco Splendid Home 1 Terminal Examination-2017 Class-VII G.KDocument2 pagesDon Bosco Splendid Home 1 Terminal Examination-2017 Class-VII G.KproodootNo ratings yet
- Valencia and Pasca, 2021 - Chromatin DynamicsDocument4 pagesValencia and Pasca, 2021 - Chromatin DynamicsJulia SCNo ratings yet
- PCR Troubleshooting-Part 1 "No Bands": Hints, Tips and Trouble Shooting For Molecular Biology TechniciansDocument6 pagesPCR Troubleshooting-Part 1 "No Bands": Hints, Tips and Trouble Shooting For Molecular Biology Technicianssam911101No ratings yet
- The Quantum Mitochondrion and Optimal HealthDocument10 pagesThe Quantum Mitochondrion and Optimal HealthmsuettmannNo ratings yet
- 1.4 Our Time On Earth2 2Document16 pages1.4 Our Time On Earth2 2aleshaNo ratings yet
- Standing - Advertisement - IITMandi - STD - 3 - 2022-Faculty - Dt. 10.10.2022Document10 pagesStanding - Advertisement - IITMandi - STD - 3 - 2022-Faculty - Dt. 10.10.2022ananda.indianrailways.clwNo ratings yet
- Exploring Biology in The Laboratory Pendarvis PDFDocument2 pagesExploring Biology in The Laboratory Pendarvis PDFJulie0% (6)
- Matta Et Al. - 2018 - Development and Characterization of A Novel LucifeDocument14 pagesMatta Et Al. - 2018 - Development and Characterization of A Novel LucifeRaghav VijNo ratings yet
- Demo LPDocument7 pagesDemo LPDaisyree DatilesNo ratings yet
- M1CHAP3Document28 pagesM1CHAP3Eilla Marie GomosNo ratings yet