You might also like
- The Hypothalamic HormoneDocument9 pagesThe Hypothalamic HormoneBanikshimulNo ratings yet
- Invertebrate Hormones: Tissue HormonesFrom EverandInvertebrate Hormones: Tissue HormonesU. S. Von EulerNo ratings yet
- IP-Worksheet-3-Endocrine System The Hypothalamic-Pituitary Axis (Estomagulang)Document2 pagesIP-Worksheet-3-Endocrine System The Hypothalamic-Pituitary Axis (Estomagulang)Filmae Estomagulang0% (1)
- The Hypothalamus and Pituitary Gland: General Principles of Endocrine DiagnosisDocument13 pagesThe Hypothalamus and Pituitary Gland: General Principles of Endocrine DiagnosisKiprono Keitany TimothyNo ratings yet
- Clinical BiochemistryDocument71 pagesClinical BiochemistryMuhamed ArsalanNo ratings yet
- HypothalamusDocument4 pagesHypothalamusLeanderVoetsNo ratings yet
- Hypothalamus and Pituitary GlandDocument39 pagesHypothalamus and Pituitary GlandGish KioiNo ratings yet
- Module 1: The Endocrine System: Learning ObjectivesDocument1 pageModule 1: The Endocrine System: Learning ObjectivesZeina SharkasNo ratings yet
- Physio 4Document4 pagesPhysio 4amjadNo ratings yet
- EN17 - Tumors of The Sellar Region - Acromegaly, Prolactinoma, Gonadotropinoma, Nonfunctioning Pituitary Adenoma, Craniopharyngioma)Document24 pagesEN17 - Tumors of The Sellar Region - Acromegaly, Prolactinoma, Gonadotropinoma, Nonfunctioning Pituitary Adenoma, Craniopharyngioma)sbobine.imsNo ratings yet
- Neurotransmitters and Psychiatry PDFDocument22 pagesNeurotransmitters and Psychiatry PDFcapriciousbelal93% (14)
- English Sistem EndocrineDocument9 pagesEnglish Sistem EndocrineNurlaili YaniNo ratings yet
- Lab Virtual Sistema Endocrino GuíaDocument17 pagesLab Virtual Sistema Endocrino Guíadavid barretoNo ratings yet
- Unit2-Hypothalamus and PituitaryDocument80 pagesUnit2-Hypothalamus and Pituitarypranutan739No ratings yet
- Cap.1 - Hypothalamic-Pituitary Sistem 2018 - 2019Document23 pagesCap.1 - Hypothalamic-Pituitary Sistem 2018 - 2019Razvan GeorgeNo ratings yet
- Functions of BodyDocument3 pagesFunctions of Bodyacouch6492No ratings yet
- New Microsoft Word DocumentDocument13 pagesNew Microsoft Word DocumentdinaNo ratings yet
- Hypothalamo Pituitary AxisDocument29 pagesHypothalamo Pituitary AxishamidNo ratings yet
- Hypothalmo-Hypophysial Axis: by Dr. Luna PhukanDocument35 pagesHypothalmo-Hypophysial Axis: by Dr. Luna PhukanAmmar QuasmiNo ratings yet
- Clinical Chemistry - PITUITARY HORMONES TRANSDocument7 pagesClinical Chemistry - PITUITARY HORMONES TRANSCamella Beatrice Lujan ValleNo ratings yet
- Endocrine System: Citation NeededDocument8 pagesEndocrine System: Citation NeededAsif Hassan BiswasNo ratings yet
- Pcol Lec Book FinalsDocument61 pagesPcol Lec Book FinalsMarco Sta AnaNo ratings yet
- The Endocrine SystemDocument101 pagesThe Endocrine SystemLeônidas ZazelisNo ratings yet
- Endocrine System: Navigation SearchDocument9 pagesEndocrine System: Navigation SearchAnnagella De JesusNo ratings yet
- Endocrinol Pred 02Document7 pagesEndocrinol Pred 02api-3706483No ratings yet
- Inbound 8692870994275491539Document6 pagesInbound 8692870994275491539MUHAMMAD ISHAQNo ratings yet
- Overview of Endo Physiology - HandoutDocument19 pagesOverview of Endo Physiology - HandoutVDiesel99No ratings yet
- Week 5&6 Endocrine SystemDocument34 pagesWeek 5&6 Endocrine Systemk.shamim1999No ratings yet
- 9F. Endokrin 2Document53 pages9F. Endokrin 2Irfan Maulana AjiNo ratings yet
- Endocrine SystemDocument14 pagesEndocrine SystemEdsel Ian S. FuentesNo ratings yet
- Endokrin MW DitransletDocument7 pagesEndokrin MW DitransletSanta SaintNo ratings yet
- LO Endocrine 1.2 Dapots 1. Hormone Tubuh in GeneralDocument3 pagesLO Endocrine 1.2 Dapots 1. Hormone Tubuh in GeneralDapot SianiparNo ratings yet
- Pituitary GlandDocument15 pagesPituitary GlandPutri Agustin Mere100% (1)
- Reproductive MedicineDocument330 pagesReproductive MedicineunknownxemNo ratings yet
- Endocrinology Notes: Veterinary PhysiologyDocument16 pagesEndocrinology Notes: Veterinary PhysiologyBrian AllanNo ratings yet
- Hypothalamus and Thalamus .PhysiologyDocument33 pagesHypothalamus and Thalamus .Physiologybareqgassan100% (1)
- Hypothalamus Pituitary Glands Adrenal CortexDocument2 pagesHypothalamus Pituitary Glands Adrenal CortexEmNo ratings yet
- Hypothalamus - Hypophysis Functional AnatomyDocument11 pagesHypothalamus - Hypophysis Functional AnatomyRajesh ReddyNo ratings yet
- tmp5B80 TMPDocument27 pagestmp5B80 TMPFrontiersNo ratings yet
- tmp4946 TMPDocument27 pagestmp4946 TMPFrontiersNo ratings yet
- Tmp3a76 TMPDocument27 pagesTmp3a76 TMPFrontiersNo ratings yet
- 3.0 Control of Anterior Pituitary SecretionDocument73 pages3.0 Control of Anterior Pituitary SecretionHomeground entertainmentNo ratings yet
- Eje H-H-O InglesDocument32 pagesEje H-H-O Inglesrafael martinezNo ratings yet
- Pituitary Gland (Hypophysis)Document28 pagesPituitary Gland (Hypophysis)Nastase Daniela EcaterinaNo ratings yet
- Pitutary GlandDocument3 pagesPitutary GlandMariannNo ratings yet
- FibromialgiaDocument7 pagesFibromialgiamatheus galvãoNo ratings yet
- Pituitary Gland - Structure and Function by Dr. Shushma KumariDocument6 pagesPituitary Gland - Structure and Function by Dr. Shushma Kumariakshdeep1747No ratings yet
- Drugs Affecting The Endocrine SystemDocument43 pagesDrugs Affecting The Endocrine SystemJustine Vens G. AgustinNo ratings yet
- Polytechnic University of The Philippines Sta. Mesa, Manila: Health Problems and BehaviorDocument9 pagesPolytechnic University of The Philippines Sta. Mesa, Manila: Health Problems and BehaviorJasmine Manzano DicalengNo ratings yet
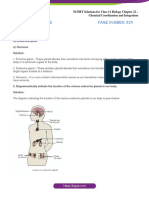
- Ncert Solutions June6 For Class 11 Biology Chapter 22 Chemical Coordination and IntegrationDocument8 pagesNcert Solutions June6 For Class 11 Biology Chapter 22 Chemical Coordination and Integrationginnisingh524No ratings yet
- Phys 6.1 and 6.2 Ant Pituitary GH and Post Pituitary NOTESDocument7 pagesPhys 6.1 and 6.2 Ant Pituitary GH and Post Pituitary NOTESsine summsun PradhanNo ratings yet
- Term Paper : Isabela State UniversityDocument9 pagesTerm Paper : Isabela State UniversityFlor SagnipNo ratings yet
- Cushing SyndromeDocument66 pagesCushing SyndromeAndi Agung RiatmojoNo ratings yet
- 4.0 The Pituitary GlandDocument68 pages4.0 The Pituitary GlandHomeground entertainmentNo ratings yet
- 2-Endocrine (Hypothalamo-Pituitary Axis and Regulatory Mechanisms) Med 2020Document29 pages2-Endocrine (Hypothalamo-Pituitary Axis and Regulatory Mechanisms) Med 2020BHUWAN BASKOTANo ratings yet
- Hypothalamic Regulation of Hormone SecretionDocument3 pagesHypothalamic Regulation of Hormone SecretionDr Sanjeeb Kumar Dey BaidyaNo ratings yet
- European Journal of Endocrinology) Corticotropin-Releasing Hormone PhysiologyDocument6 pagesEuropean Journal of Endocrinology) Corticotropin-Releasing Hormone PhysiologyBenedictus Yudha BaskaraNo ratings yet
- Pituitary HormonesDocument75 pagesPituitary HormonesRinkiNo ratings yet
- 5 Axe HH Anglais VioletDocument29 pages5 Axe HH Anglais Violetyoyo mumbaNo ratings yet
- Pnle Medical Surgical ReviewDocument17 pagesPnle Medical Surgical ReviewKyla Rose ToledoNo ratings yet
- Chapter 11Document11 pagesChapter 11christian anchetaNo ratings yet
- N-Cadherin and Muller GliaDocument12 pagesN-Cadherin and Muller GliaRebeca VictoriaNo ratings yet
- Saliva: DR - Deepshikha Soni Mds 1 YearDocument90 pagesSaliva: DR - Deepshikha Soni Mds 1 Yeardisha 146jandialNo ratings yet
- Dermatopathology Diagnosis by First Impression 3rd Edition Ebook PDFDocument57 pagesDermatopathology Diagnosis by First Impression 3rd Edition Ebook PDFmario.becker252100% (42)
- Anatomy N PhysiologyDocument10 pagesAnatomy N Physiologynaem nadzriNo ratings yet
- Leishmaniasis: Leishmania Donovani (Complex)Document32 pagesLeishmaniasis: Leishmania Donovani (Complex)javeria choudharyNo ratings yet
- Science MCQDocument45 pagesScience MCQGeneric nameNo ratings yet
- Enzyme Practice Quiz Questions and AnswersDocument11 pagesEnzyme Practice Quiz Questions and AnswersMa Angelica Shane Navarro100% (1)
- Progress in Neuropsychopharmacology & Biological PsychiatryDocument8 pagesProgress in Neuropsychopharmacology & Biological PsychiatrySumedhaNo ratings yet
- ELEC4810 Notes-1 PDFDocument71 pagesELEC4810 Notes-1 PDFKwan ChanNo ratings yet
- Aerobic and Anaerobic RespirationDocument6 pagesAerobic and Anaerobic Respiration10A 27 Mahek PanirwalaNo ratings yet
- Science 9: Remember My PartsDocument6 pagesScience 9: Remember My PartsJan Matthew NuezNo ratings yet
- Nervous SystemDocument48 pagesNervous SystemSuvalari Mimi JonathanNo ratings yet
- The Group Actions of MuscleDocument21 pagesThe Group Actions of MuscleMuhammad Owais qarniNo ratings yet
- ANPH Midterm NotesDocument44 pagesANPH Midterm NotesClaudine Jo B. TalabocNo ratings yet
- Monday 15 June 2020: Biology BDocument36 pagesMonday 15 June 2020: Biology Bbody fayezNo ratings yet
- Micro - Rejection of Kidney TransplantDocument28 pagesMicro - Rejection of Kidney TransplantMahmoud hilmyNo ratings yet
- CytokinesDocument2 pagesCytokinesCesar RodrìguezNo ratings yet
- (S-W5-Sun-Gen.S) (By Dr. Emad) Gall Bladder 1Document28 pages(S-W5-Sun-Gen.S) (By Dr. Emad) Gall Bladder 1Haider Nadhem AL-rubaiNo ratings yet
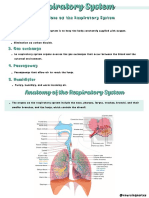
- Respiratory SystemDocument12 pagesRespiratory SystemTricia CabiliNo ratings yet
- DM-1 - Lesson 18Document9 pagesDM-1 - Lesson 18Amit GodaraNo ratings yet
- DALLALhead Nurse AutosavedDocument77 pagesDALLALhead Nurse AutosavedmulanNo ratings yet
- ازمایشDocument3 pagesازمایشv.shaakaramiNo ratings yet
- Grade 6 CirculatoryDocument32 pagesGrade 6 Circulatoryalvarez9654No ratings yet
- Risk NCP Decreased Cardiac OutputDocument2 pagesRisk NCP Decreased Cardiac OutputMICHELLE FACTONo ratings yet
- Cap DependentDocument6 pagesCap DependentKAIST이박터No ratings yet
- Negative and Positive FeedbackDocument12 pagesNegative and Positive Feedbackzshia shang limNo ratings yet
- HaptenDocument3 pagesHaptenThu Trang TranNo ratings yet
- Biology For The IB Diploma - Option A Neurobiology and BehaviourDocument51 pagesBiology For The IB Diploma - Option A Neurobiology and BehaviourjjkjljNo ratings yet
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4.5 out of 5 stars4.5/5 (6)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (598)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseFrom EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseRating: 4.5 out of 5 stars4.5/5 (52)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (217)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessFrom EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessRating: 4 out of 5 stars4/5 (18)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (111)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceFrom EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceRating: 4 out of 5 stars4/5 (633)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Minds Make Societies: How Cognition Explains the World Humans CreateFrom EverandMinds Make Societies: How Cognition Explains the World Humans CreateRating: 4.5 out of 5 stars4.5/5 (24)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Remnants of Ancient Life: The New Science of Old FossilsFrom EverandRemnants of Ancient Life: The New Science of Old FossilsRating: 3 out of 5 stars3/5 (3)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)