You might also like
- Scientific Report On Transport Mechanism: Ultrafiltration Is A Special Type ofDocument3 pagesScientific Report On Transport Mechanism: Ultrafiltration Is A Special Type ofBrixNo ratings yet
- A New Theory of Amoeboid Locomotion: Y. Theoret. Biol. (1963) 4, 124-141Document18 pagesA New Theory of Amoeboid Locomotion: Y. Theoret. Biol. (1963) 4, 124-141graviphotonNo ratings yet
- Essential Idea: Structure and Function Are Correlated in The Phloem of PlantsDocument14 pagesEssential Idea: Structure and Function Are Correlated in The Phloem of PlantsINDRANI GOSWAMINo ratings yet
- Microswimming in Viscoelastic FluidsDocument16 pagesMicroswimming in Viscoelastic FluidsMuhammad MusharafNo ratings yet
- Congestive Cardiac FalureDocument57 pagesCongestive Cardiac FalurelightningiceNo ratings yet
- Myosin Motors Drive Long Range AlignmentDocument12 pagesMyosin Motors Drive Long Range AlignmentAlexNo ratings yet
- Velez 2013Document14 pagesVelez 2013jucasegaNo ratings yet
- Fluid Circulation Driven by Collectively Organized Metachronal Waves in Swimming T. Aceiti NematodesDocument13 pagesFluid Circulation Driven by Collectively Organized Metachronal Waves in Swimming T. Aceiti NematodesqgravNo ratings yet
- Wadhwa Et Al-2021-Nature Reviews Microbiology-1Document13 pagesWadhwa Et Al-2021-Nature Reviews Microbiology-1pridNo ratings yet
- The Lymphatic Fluid: Laura SantambrogioDocument23 pagesThe Lymphatic Fluid: Laura SantambrogioktorooNo ratings yet
- 1 s2.0 S2211379717312421 MainDocument8 pages1 s2.0 S2211379717312421 MainAdil IhsanNo ratings yet
- Full Dynamics of A Red Blood Cell in Shear Ow: 20808 - 20813 Pnas December 18, 2012 Vol. 109 No. 51Document6 pagesFull Dynamics of A Red Blood Cell in Shear Ow: 20808 - 20813 Pnas December 18, 2012 Vol. 109 No. 51rezqNo ratings yet
- Traffic - 2004 - Etienne Manneville - Actin and Microtubules in Cell Motility Which One Is in ControlDocument8 pagesTraffic - 2004 - Etienne Manneville - Actin and Microtubules in Cell Motility Which One Is in ControlPinaki NayakNo ratings yet
- Modeling of Spontaneous Penetration of Viscoelastic FluidsDocument10 pagesModeling of Spontaneous Penetration of Viscoelastic Fluidssu67983629No ratings yet
- Role of CFTR in Epithelial Physiology: Cellular and Molecular Life SciencesDocument23 pagesRole of CFTR in Epithelial Physiology: Cellular and Molecular Life Sciencesshaheena ahmedNo ratings yet
- 824-Texto Del Manuscrito Completo (Cuadros y Figuras Insertos) - 4381-1!10!20120828Document6 pages824-Texto Del Manuscrito Completo (Cuadros y Figuras Insertos) - 4381-1!10!20120828Tallie ZeidlerNo ratings yet
- Pulsing Corals: A Story of Scale and Mixing: Original ArticleDocument14 pagesPulsing Corals: A Story of Scale and Mixing: Original ArticleWyatt GrayNo ratings yet
- Understanding Trabecular Meshwork Physiology: A Key To The Control of Intraocular Pressure?Document5 pagesUnderstanding Trabecular Meshwork Physiology: A Key To The Control of Intraocular Pressure?RaissaNo ratings yet
- Cho2016 Article RolesOfCross-MembraneTransportDocument13 pagesCho2016 Article RolesOfCross-MembraneTransportTaurus PearsonNo ratings yet
- 3 The Electrochemical Basis of Nerve FunctionDocument33 pages3 The Electrochemical Basis of Nerve FunctionEvets JarusNo ratings yet
- A New Link Between The Retrograde Actin Flow and FDocument11 pagesA New Link Between The Retrograde Actin Flow and FPinaki NayakNo ratings yet
- Physiology of Cerebrospinal Fluid Circulation: ReviewDocument8 pagesPhysiology of Cerebrospinal Fluid Circulation: Reviewnataly yañezNo ratings yet
- Living Liquid Crystals: SignificanceDocument6 pagesLiving Liquid Crystals: SignificanceJonathan PatelNo ratings yet
- Wiig 2012Document56 pagesWiig 2012SDNKSJNDJKNo ratings yet
- DK2823 CH02Document46 pagesDK2823 CH02Anis WidiarsihNo ratings yet
- Vacuolar Control of Stomatal Opening Revealed by 3D Imaging of The Guard CellsDocument10 pagesVacuolar Control of Stomatal Opening Revealed by 3D Imaging of The Guard CellsPunta IndratomoNo ratings yet
- Anatomy and Physiology - The Unity of Form and Function 5th Ed - Saladin (McGraw-Hill, 2009) BBSDocument4 pagesAnatomy and Physiology - The Unity of Form and Function 5th Ed - Saladin (McGraw-Hill, 2009) BBSdixama9519No ratings yet
- 4 Circulatory SystemDocument23 pages4 Circulatory SystemSundari D'yojapa NonemNo ratings yet
- PHY Fluids WBC KineticsDocument22 pagesPHY Fluids WBC KineticsRamesh KavaliNo ratings yet
- Human Anatomy 4th Edition Saladin Solutions ManualDocument25 pagesHuman Anatomy 4th Edition Saladin Solutions ManualLeslieBuchanannjfy100% (52)
- Full Download Human Anatomy 4th Edition Saladin Solutions ManualDocument35 pagesFull Download Human Anatomy 4th Edition Saladin Solutions Manualvocality.duettoqw603100% (36)
- Hydrogel 5Document17 pagesHydrogel 5Rana Sabouni TabariNo ratings yet
- Darrow1950 PDFDocument9 pagesDarrow1950 PDFLuís Eduardo CruvinelNo ratings yet
- Internship PHD Physarum Durand 2018 ICFPDocument1 pageInternship PHD Physarum Durand 2018 ICFPje_suis_anonymeNo ratings yet
- PERvsResonanceJellyfishPaper FINALDocument32 pagesPERvsResonanceJellyfishPaper FINALMarcusNo ratings yet
- Brugues PNAS 2010Document10 pagesBrugues PNAS 2010Pierre NassoyNo ratings yet
- Containing The Cell: Cell Membrane: Part I: Locating Physiology On The Web of KnowledgeDocument5 pagesContaining The Cell: Cell Membrane: Part I: Locating Physiology On The Web of KnowledgeEly TrajicoNo ratings yet
- Transport CirculationDocument39 pagesTransport CirculationClara MaeNo ratings yet
- Living Liquid Crystals: Shuang Zhou, Andrey Sokolov, Oleg D Lavrentovich, Igor S AransonDocument32 pagesLiving Liquid Crystals: Shuang Zhou, Andrey Sokolov, Oleg D Lavrentovich, Igor S AransonTyler Jordan DavisonNo ratings yet
- Swimming by Reciprocal Motion at Low Reynolds Number: ArticleDocument8 pagesSwimming by Reciprocal Motion at Low Reynolds Number: Articlejuan felipeNo ratings yet
- Anatomy and Pa Tho Physiology of Aqueous Production and OutflowDocument34 pagesAnatomy and Pa Tho Physiology of Aqueous Production and OutflowChristine TanNo ratings yet
- Discuss Physiology of Aqueous Formation and Its Drainage and Relate It To How Intraocular Pressure Is ControlledDocument8 pagesDiscuss Physiology of Aqueous Formation and Its Drainage and Relate It To How Intraocular Pressure Is ControlledPrince ChipunguNo ratings yet
- 5-Circulatory System Open Vs ClosedDocument9 pages5-Circulatory System Open Vs Closedchristina8reales8ca8No ratings yet
- Superconducting Liquid Crystalline Meridians-LibreDocument16 pagesSuperconducting Liquid Crystalline Meridians-LibreAmentiHall100% (2)
- A Computer Model of The Lymphatic System PDFDocument17 pagesA Computer Model of The Lymphatic System PDFnnimrodoNo ratings yet
- Rotational DynamicsDocument6 pagesRotational DynamicsjucasegaNo ratings yet
- Bir 1987 24606Document7 pagesBir 1987 24606Constanza Martinez ArancibiaNo ratings yet
- Contemporary PhysicsDocument26 pagesContemporary PhysicsIffah KhairunnisaNo ratings yet
- Cold Spring Harb Perspect Biol-2017-Okamoto-cshperspect.a027839Document21 pagesCold Spring Harb Perspect Biol-2017-Okamoto-cshperspect.a027839crysraldixxNo ratings yet
- (People and Ideas) Daniel C. Tosteson (Auth.), Daniel C. Tosteson (Eds.) - Membrane Transport - People and Ideas (1989, Springer New York)Document410 pages(People and Ideas) Daniel C. Tosteson (Auth.), Daniel C. Tosteson (Eds.) - Membrane Transport - People and Ideas (1989, Springer New York)gmansh90No ratings yet
- Anatomyofthevestibularsystem Areview PDFDocument8 pagesAnatomyofthevestibularsystem Areview PDFJacqueline Rebolledo BravoNo ratings yet
- Weak Synchronization and Large-Scale Collective OscillationDocument5 pagesWeak Synchronization and Large-Scale Collective OscillationTrung PhanNo ratings yet
- Journal FreeDocument36 pagesJournal FreeRiccardo Spinoza SilvaNo ratings yet
- Fluid Flows and Forces in Development Functions, Features and Biophysical PrinciplesDocument19 pagesFluid Flows and Forces in Development Functions, Features and Biophysical PrinciplesSantiago ToroNo ratings yet
- Algal Flagella PDFDocument6 pagesAlgal Flagella PDFmanoj_rkl_07No ratings yet
- Inter 2nd Year Botany 1.transport - Plants Study Material PDFDocument8 pagesInter 2nd Year Botany 1.transport - Plants Study Material PDFsingoj.bhargava chary100% (10)
- Structure: As As AsDocument32 pagesStructure: As As AsSuchitra SharmaNo ratings yet
- CambioDocument16 pagesCambioEdson AugustoNo ratings yet
- Transport in HumansDocument1 pageTransport in Humansapi-3714421100% (1)
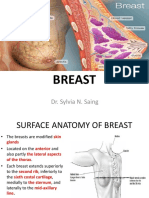
- Breast: Dr. Sylvia N. SaingDocument26 pagesBreast: Dr. Sylvia N. SaingRichard 151289100% (1)
- Dictyosome: Modifying, and Packaging Proteins and Lipids Into Vesicles ForDocument11 pagesDictyosome: Modifying, and Packaging Proteins and Lipids Into Vesicles ForASHFAQ AHMAD100% (1)
- Connective TissueDocument62 pagesConnective Tissuehnm mnhNo ratings yet
- Chapter 26 Guyton and HallDocument5 pagesChapter 26 Guyton and HallSanna Asila AkramNo ratings yet
- Anatomy and Physiology of The Circulatory SystemDocument7 pagesAnatomy and Physiology of The Circulatory SystemAlessandro Arellano HmuaNo ratings yet
- Female Reproductive SystemDocument7 pagesFemale Reproductive SystemKomail SrnNo ratings yet
- Diamond-Blackfan AnemiaDocument5 pagesDiamond-Blackfan AnemiaTalal 197No ratings yet
- Life Flows Because of You..Document40 pagesLife Flows Because of You..Klemen JamnikNo ratings yet
- Neuroscience: Science of The Brain in NepaliDocument61 pagesNeuroscience: Science of The Brain in NepaliInternational Brain Research Organization100% (1)
- BrainEdge IAS Mains SyllabusDocument153 pagesBrainEdge IAS Mains SyllabusAnantjit Singh SahniNo ratings yet
- Urinary SystemDocument3 pagesUrinary SystemFadhil Hussam AhmedNo ratings yet
- Presbycusis: Dr. George Backleh Dept. of Otolaryngology/ Head and Neck Surgery Hadassah University Hospital JerusalemDocument16 pagesPresbycusis: Dr. George Backleh Dept. of Otolaryngology/ Head and Neck Surgery Hadassah University Hospital Jerusalemrezky_oktarianti_syahputriNo ratings yet
- Beaconhouse School System: Biology ConceptsDocument3 pagesBeaconhouse School System: Biology ConceptsUsman AmeenNo ratings yet
- BIO201L Lab 5 TissuesDocument12 pagesBIO201L Lab 5 Tissuesmyra Thiong'oNo ratings yet
- Exercise 1Document35 pagesExercise 1Van Labasano67% (3)
- Chapter 2 - Anatomy Posture and Body MechanicsDocument53 pagesChapter 2 - Anatomy Posture and Body MechanicsRogini Sindhu100% (6)
- Muscular System Study Guide 2015 AnswersDocument16 pagesMuscular System Study Guide 2015 AnswersLinet HuchuNo ratings yet
- CHAPTER Respiration and CirculationDocument5 pagesCHAPTER Respiration and CirculationlatasabarikNo ratings yet
- Urinary System PDFDocument27 pagesUrinary System PDFfiona100% (3)
- Acute GastroenteritisDocument56 pagesAcute Gastroenteritisneil052298% (46)
- 6th Class Digestive SystemDocument20 pages6th Class Digestive SystemJuvy MarquinezNo ratings yet
- Histology Full McqsDocument157 pagesHistology Full McqsElijah Kamani100% (1)
- Project in BiologyDocument5 pagesProject in BiologyViah TronzalNo ratings yet
- Cell JunctionsDocument40 pagesCell JunctionsAhmad MursyidNo ratings yet
- Bowel Obstruction Case StudyDocument11 pagesBowel Obstruction Case StudyFatimah Batool NawazNo ratings yet
- Mindmap Bio621 Chapter1Document3 pagesMindmap Bio621 Chapter1MizahNo ratings yet
- PhysioEx Exercise 11 Activity 4 - MD3N3Document4 pagesPhysioEx Exercise 11 Activity 4 - MD3N3Dorlisca Montes GarcesNo ratings yet
- Lecture2 GranulopoiesisDocument9 pagesLecture2 GranulopoiesisAfifa Prima GittaNo ratings yet
- Dysphagia Project - Literature ReviewDocument26 pagesDysphagia Project - Literature ReviewJeerunnun UparipahkNo ratings yet