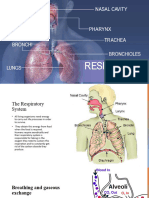
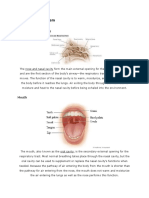
You might also like
- Respiratory gymnastics (Translated): Purification - Health - Strength - EnergyFrom EverandRespiratory gymnastics (Translated): Purification - Health - Strength - EnergyNo ratings yet
- The Effect of Exercise On The Cardiorespiratory SystemDocument15 pagesThe Effect of Exercise On The Cardiorespiratory SystemBASAVARAJ100% (1)
- The Casket of MedicineDocument104 pagesThe Casket of Medicinejivasumana100% (4)
- Method Statement For Installation & Insulation of Buried Chilled Water PipingDocument8 pagesMethod Statement For Installation & Insulation of Buried Chilled Water PipingBinod DavisNo ratings yet
- Atwood - 1984 - Molten Salt TechnologyDocument536 pagesAtwood - 1984 - Molten Salt TechnologyLanly Romuel100% (1)
- Respiratory SystemDocument32 pagesRespiratory Systemapi-202513022No ratings yet
- CQ Brochure - English A4Document20 pagesCQ Brochure - English A4mrbookaniNo ratings yet
- Lesson 3: Gas ExchangeDocument7 pagesLesson 3: Gas ExchangeChenul WickramasingheNo ratings yet
- Human Body Book | Introduction to the Respiratory System | Children's Anatomy & Physiology EditionFrom EverandHuman Body Book | Introduction to the Respiratory System | Children's Anatomy & Physiology EditionNo ratings yet
- Gas Exchange in AnimalsDocument20 pagesGas Exchange in AnimalsMaria Theresa HerreroNo ratings yet
- Chapter II - Respiratory SystemDocument3 pagesChapter II - Respiratory SystemIndranil SinhaNo ratings yet
- Tank Heating DiscussionsDocument26 pagesTank Heating DiscussionsTHERMAX007No ratings yet
- Gaseous ExchangeDocument37 pagesGaseous ExchangeSolángé MirembeNo ratings yet
- Anatomy and Physiology of The Respiratory SystemDocument5 pagesAnatomy and Physiology of The Respiratory SystemxoxosvwNo ratings yet
- Ventury Scrubber Design MethodDocument3 pagesVentury Scrubber Design MethodPukhraj DagaNo ratings yet
- Chemical Industry Pakistan 2030 PDFDocument151 pagesChemical Industry Pakistan 2030 PDFhatim75% (8)
- Essay 1Document2 pagesEssay 1Ni IluhNo ratings yet
- M132 Respiratory 2022Document28 pagesM132 Respiratory 2022Bana ItanNo ratings yet
- Chapter 12Document28 pagesChapter 12براءة أحمد السلاماتNo ratings yet
- Chapter 12Document28 pagesChapter 12Ammar SmadiNo ratings yet
- 3 1Document7 pages3 1Zainab FahadNo ratings yet
- Chapter 11 BioDocument18 pagesChapter 11 BioRylee LNo ratings yet
- BIO 2701 Lecture Notes Exchange With The EnvironmentDocument11 pagesBIO 2701 Lecture Notes Exchange With The EnvironmentDorothy MayakaNo ratings yet
- Diffusion and Human Respiratory SystemDocument9 pagesDiffusion and Human Respiratory SystemPriscilla AshwiniNo ratings yet
- OCR (A) Biology A-Level: Topic 3.1: Exchange and TransportDocument4 pagesOCR (A) Biology A-Level: Topic 3.1: Exchange and TransportNguyen Thi Thien MinhNo ratings yet
- Breathing and Exchange of Gases-HumansDocument14 pagesBreathing and Exchange of Gases-Humansnihalbolla2023No ratings yet
- Breathing and Gas ExchangeDocument8 pagesBreathing and Gas ExchangeSumaiya AnwarNo ratings yet
- CH 11 Gas Exchange in HumansDocument4 pagesCH 11 Gas Exchange in HumansPranitha RaviNo ratings yet
- Gas ExchangeDocument6 pagesGas Exchangerasha nadaNo ratings yet
- 12.respiratory SystemDocument52 pages12.respiratory SystemRulani PrinceNo ratings yet
- Gas Exchange NewDocument30 pagesGas Exchange Newmariaamkhaann22No ratings yet
- 1.4 Lungs & DiseasesDocument10 pages1.4 Lungs & DiseasesRayonesh RayanaNo ratings yet
- Gaseous Exchange in The Mammalian Lungs: by Vytautas KontrimasDocument10 pagesGaseous Exchange in The Mammalian Lungs: by Vytautas KontrimascluendoNo ratings yet
- Gas Exchange in HumansDocument23 pagesGas Exchange in HumansVNo ratings yet
- Science 9 1.1 LessonDocument25 pagesScience 9 1.1 LessonHdisbdbd NfjdnbdncNo ratings yet
- Gas Exchange in The LungsDocument30 pagesGas Exchange in The LungsYasasviNo ratings yet
- Thought For The Week: Gossips & Jealousy Bring: - Subtraction of Love - Division of FriendshipDocument74 pagesThought For The Week: Gossips & Jealousy Bring: - Subtraction of Love - Division of FriendshipSepi SammyNo ratings yet
- Gas Exchange System SAT ContentDocument3 pagesGas Exchange System SAT ContentJohn Osborne100% (1)
- Features of Gas Exchange Surfaces: The Alveolus Is The Gas Exchange Surface in HumansDocument9 pagesFeatures of Gas Exchange Surfaces: The Alveolus Is The Gas Exchange Surface in HumansM. ShayanNo ratings yet
- Respiratory SystemDocument12 pagesRespiratory Systemab4943409No ratings yet
- Respiratory SystemDocument32 pagesRespiratory SystemvhieeelgbmNo ratings yet
- Module 2Document40 pagesModule 2Sharanya BasakNo ratings yet
- Reflection Respiratory SystemDocument4 pagesReflection Respiratory SystemAin Sufiza0% (1)
- Respiratory SystemDocument12 pagesRespiratory SystemM. Farkhan HabibNo ratings yet
- Achievement: Gas Exchange in Animals InsectsDocument8 pagesAchievement: Gas Exchange in Animals Insectssimon chanNo ratings yet
- Bio Notes Respiration in AnimalsDocument3 pagesBio Notes Respiration in AnimalssamanthahuangsamanthaNo ratings yet
- Lecture 4 THE RESPIRATORY SYSTEMDocument48 pagesLecture 4 THE RESPIRATORY SYSTEMZara16No ratings yet
- Breathing and RespirationDocument30 pagesBreathing and Respirationhamzahazard527No ratings yet
- Breathing and Gaseous Exchnage by Sherrie-Ann Wilson-BrownDocument85 pagesBreathing and Gaseous Exchnage by Sherrie-Ann Wilson-BrownNayoka EnnisNo ratings yet
- The Respiratory SystemDocument2 pagesThe Respiratory SystemconnievelardeNo ratings yet
- Summary Notes For IGCSE BiologyDocument3 pagesSummary Notes For IGCSE BiologyStudy GuyNo ratings yet
- Respiratory and Circulatory SystemDocument10 pagesRespiratory and Circulatory Systemainabukhari20No ratings yet
- Breathing and Exchange of GasesDocument8 pagesBreathing and Exchange of GasesgoswamiuwuNo ratings yet
- The Respiratory System and RespirationDocument7 pagesThe Respiratory System and RespirationAntonateNo ratings yet
- 6.4 QuestionsDocument18 pages6.4 QuestionsAurelija GardžiulytėNo ratings yet
- s.4 Biology Notes Gaseous Exchange and RespirationDocument14 pagess.4 Biology Notes Gaseous Exchange and RespirationRonald RomNo ratings yet
- Chapter10. Class 10th (Page7)Document7 pagesChapter10. Class 10th (Page7)Abdus samiNo ratings yet
- bio 统一小考-chapter 8Document8 pagesbio 统一小考-chapter 822 Lee Xin Yong 李芯瑢No ratings yet
- Dr. Sunita Saxena - Breathing and Exchange of GassesDocument48 pagesDr. Sunita Saxena - Breathing and Exchange of GassesDivya AgarawalNo ratings yet
- Respiratory SystemDocument14 pagesRespiratory SystemTedi FetoNo ratings yet
- Gaseous Exchange A LevelDocument33 pagesGaseous Exchange A LevelemmanuelkeetoNo ratings yet
- The Respiratory System-11 By:-Dr. Hazim AL-RawiDocument12 pagesThe Respiratory System-11 By:-Dr. Hazim AL-RawiDrAli Al-FendiNo ratings yet
- Study Materials: Vedantu Innovations Pvt. Ltd. Score High With A Personal Teacher, Learn LIVE Online!Document17 pagesStudy Materials: Vedantu Innovations Pvt. Ltd. Score High With A Personal Teacher, Learn LIVE Online!Saraswati PNo ratings yet
- Respiratory SystemDocument6 pagesRespiratory SystemCraigNo ratings yet
- Respiratory System: Nose and Nasal CavityDocument8 pagesRespiratory System: Nose and Nasal CavityJasper AlbuferaNo ratings yet
- Respiratory SystemDocument33 pagesRespiratory Systemamulyajadhav119No ratings yet
- Respiratory System-1Document84 pagesRespiratory System-1pranjal raiNo ratings yet
- How Do Humans Breathe? Science Book Age 8 | Children's Biology BooksFrom EverandHow Do Humans Breathe? Science Book Age 8 | Children's Biology BooksNo ratings yet
- Nickel Molybdate Catalysts and Their Use in The Selective Oxidation of HydrocarbonsDocument60 pagesNickel Molybdate Catalysts and Their Use in The Selective Oxidation of HydrocarbonsSandy MartinezNo ratings yet
- NQ 34746Document208 pagesNQ 34746bittu kumarNo ratings yet
- First Estimate of Δ θ (0) : 1 2 cr crDocument41 pagesFirst Estimate of Δ θ (0) : 1 2 cr crFNo ratings yet
- Alcohol De-Addiction Ayurvedic TherapyDocument4 pagesAlcohol De-Addiction Ayurvedic TherapyklcmnNo ratings yet
- Viscosity Cup PDFDocument16 pagesViscosity Cup PDFSaut Maruli Tua SamosirNo ratings yet
- GROUP II Molecular Bio PresentationDocument20 pagesGROUP II Molecular Bio PresentationObed MuhweziNo ratings yet
- Spe 200369 PaDocument13 pagesSpe 200369 PaYousif IraqiNo ratings yet
- Biofertilizer Technology ManualDocument39 pagesBiofertilizer Technology ManualneharockNo ratings yet
- The Reaction of Metals With Acids: TopicDocument3 pagesThe Reaction of Metals With Acids: TopicHingryd RauenNo ratings yet
- Effluent TreatmentDocument21 pagesEffluent TreatmentRadhika AnandkumarNo ratings yet
- Package Insert - PYROSPERSE™ Dispersing Agent (English) - Original - 27876Document7 pagesPackage Insert - PYROSPERSE™ Dispersing Agent (English) - Original - 27876Putri DozanNo ratings yet
- Chapter 14Document29 pagesChapter 14Ella KabongoNo ratings yet
- How To Revive Cultures - Atcc GuideDocument3 pagesHow To Revive Cultures - Atcc GuidesyriusNo ratings yet
- UNIT IOG SAMPLE MATERIAL - Hydrocarbon Process Safety PDFDocument7 pagesUNIT IOG SAMPLE MATERIAL - Hydrocarbon Process Safety PDFBakiNo ratings yet
- Exploration - Phase ChangesDocument6 pagesExploration - Phase ChangesCARYS BROWNNo ratings yet
- Community Herbal Monograph On Devils ClawDocument7 pagesCommunity Herbal Monograph On Devils ClawLesley HollardNo ratings yet
- Abst PDFDocument167 pagesAbst PDFKrishnamoorthi BalasubaramanianNo ratings yet
- Graphite Powder 500GDocument7 pagesGraphite Powder 500Gakbar macheteNo ratings yet
- (4130) Resonet Sample Paper Moving To Class 9thDocument10 pages(4130) Resonet Sample Paper Moving To Class 9thShubhamNo ratings yet
- Comparison of The Boiling Water of Bay Leaves (Eugenia: Polyantha Wight) CONCENTRATION OF 100%, 70%, 50% AND 1%Document4 pagesComparison of The Boiling Water of Bay Leaves (Eugenia: Polyantha Wight) CONCENTRATION OF 100%, 70%, 50% AND 1%MadeRhamaNo ratings yet
- Effect of Varying Mixing & Compaction Temperature On Marshall Properties of Bituminous Concrete MixDocument44 pagesEffect of Varying Mixing & Compaction Temperature On Marshall Properties of Bituminous Concrete Mixshiva5059100% (1)
- New Doc 4Document46 pagesNew Doc 4imnithin100% (1)