You might also like
- L9 PDFDocument22 pagesL9 PDFMiles HuiNo ratings yet
- Front Page: Effects of Systemic Vascular Resistance On The BodyDocument1 pageFront Page: Effects of Systemic Vascular Resistance On The BodyRodel Aguila SañoNo ratings yet
- Venous Return in Critical IllnessDocument15 pagesVenous Return in Critical IllnessInês MendonçaNo ratings yet
- School of Biomechanic NSaaaaDocument30 pagesSchool of Biomechanic NSaaaaRadojica MaliNo ratings yet
- Coronary Blood Flow Regulation and DeterminantsDocument4 pagesCoronary Blood Flow Regulation and Determinantsjordandenise2No ratings yet
- Special Circulation - UnpriDocument44 pagesSpecial Circulation - UnpriAnnisNo ratings yet
- Local Control of Blood FlowDocument12 pagesLocal Control of Blood FlowMiles HuiNo ratings yet
- 40.neural (Extrinsic) Regulation of Cardiac Performance, Sympathetci and Parasympathetic Effects. Humoral Factors Affecting Cardiac PerformanceDocument3 pages40.neural (Extrinsic) Regulation of Cardiac Performance, Sympathetci and Parasympathetic Effects. Humoral Factors Affecting Cardiac PerformanceNektarios TsakalosNo ratings yet
- Shock PresentaionDocument30 pagesShock PresentaionKennedy Ng'andweNo ratings yet
- Circulation Physiology: Dr. Dara Rosmailina Reviews Key ConceptsDocument48 pagesCirculation Physiology: Dr. Dara Rosmailina Reviews Key ConceptsYudi1223No ratings yet
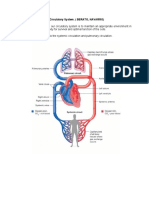
- The Circulatory System ExplainedDocument51 pagesThe Circulatory System ExplainedMohammed ahmedNo ratings yet
- Cardiac PharmocologyDocument17 pagesCardiac Pharmocologysuhas.kandeNo ratings yet
- Physiology of The Circulatory SystemDocument12 pagesPhysiology of The Circulatory SystemRugene Naragas BeratoNo ratings yet
- Liver Anatomy and PhysiologyDocument8 pagesLiver Anatomy and PhysiologyZzzemajNo ratings yet
- 23circulation Part 1Document13 pages23circulation Part 1Jaydave PatelNo ratings yet
- Blood PressureDocument58 pagesBlood PressureAyurveda PgNo ratings yet
- (v.e) Lecture 2 - Renal CirculationDocument23 pages(v.e) Lecture 2 - Renal Circulationagboolaaminat183No ratings yet
- Central Venous Pressure: Its Clinical Use and Role in Cardiovascular DynamicsFrom EverandCentral Venous Pressure: Its Clinical Use and Role in Cardiovascular DynamicsNo ratings yet
- Hepatology Research - 2007 - Lautt - Regulatory Processes Interacting To Maintain Hepatic Blood Flow Constancy VascularDocument13 pagesHepatology Research - 2007 - Lautt - Regulatory Processes Interacting To Maintain Hepatic Blood Flow Constancy VascularJosé LeãoNo ratings yet
- Blood Pressure RegulationDocument3 pagesBlood Pressure RegulationiMaibelle BelleNo ratings yet
- EDEMA - 2023-2024 - 15 ModuleDocument7 pagesEDEMA - 2023-2024 - 15 Modulerislariyas13No ratings yet
- CVS PharmacologyDocument19 pagesCVS Pharmacologymug ashNo ratings yet
- Autoregulation of Blood Flow - 210826 - 080526Document1 pageAutoregulation of Blood Flow - 210826 - 0805266qhx62pr42No ratings yet
- Mechanisms of Acute Renal Failure: C. J. L L. H C. O. S. SDocument8 pagesMechanisms of Acute Renal Failure: C. J. L L. H C. O. S. Sハルァン ファ烏山No ratings yet
- Cardio Resource UnitsDocument31 pagesCardio Resource Unitsgraveheart11No ratings yet
- Cardiovascular PhysiologyDocument5 pagesCardiovascular Physiologycentrino1750% (2)
- Relevant Equine Renal Anatomy, Physiology and Mechanisms of AKI ReviewDocument12 pagesRelevant Equine Renal Anatomy, Physiology and Mechanisms of AKI ReviewMarilú ValdepeñaNo ratings yet
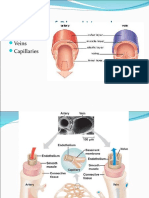
- The Cardiovascular System - Blood VesselsDocument39 pagesThe Cardiovascular System - Blood VesselsMohamed YousufNo ratings yet
- Small Animal Cardiology PDFDocument208 pagesSmall Animal Cardiology PDFRoberto AmpueroNo ratings yet
- Physiological Regulation of Arterial Blood PressureDocument36 pagesPhysiological Regulation of Arterial Blood PressureFrancesNo ratings yet
- Circulation or Peripheral CirculationDocument3 pagesCirculation or Peripheral CirculationOldy MutiaraNo ratings yet
- (O. Nelson) Small Animal CardiologyDocument208 pages(O. Nelson) Small Animal CardiologyLoredana Gutu100% (1)
- Cardiovascular PharmacologyDocument20 pagesCardiovascular PharmacologyAliceAliceNo ratings yet
- Blood Pressure HomeostasisDocument7 pagesBlood Pressure HomeostasisHassan Al SinanNo ratings yet
- 25circulation Part 3Document15 pages25circulation Part 3Jaydave PatelNo ratings yet
- Pathophysiology and Etiology of Edema - IDocument9 pagesPathophysiology and Etiology of Edema - IBrandy MaddoxNo ratings yet
- Unit 3 Blood As A Transport MediumDocument43 pagesUnit 3 Blood As A Transport MediumKemoy FrancisNo ratings yet
- Lecture 6 Cardiovascular: Vascular System - The HemodynamicsDocument61 pagesLecture 6 Cardiovascular: Vascular System - The HemodynamicsAndreea ŞtefănescuNo ratings yet
- Understanding HemodynamicsDocument41 pagesUnderstanding HemodynamicsBerihunNo ratings yet
- Inotropes and Vasoconstictor PackageDocument25 pagesInotropes and Vasoconstictor PackageYoussef MokdadNo ratings yet
- CirculationDocument11 pagesCirculationRishikaphriya RauichandranNo ratings yet
- MF-local Circulation 2016Document61 pagesMF-local Circulation 2016OnSolomonNo ratings yet
- Cardiovascular Diseases and LiverDocument20 pagesCardiovascular Diseases and LiverKrishna KumarNo ratings yet
- Distribution of Blood Flow at Rest and at ExerciseDocument5 pagesDistribution of Blood Flow at Rest and at ExerciseMohammad AliNo ratings yet
- Physiology RAASDocument3 pagesPhysiology RAASAulia Mahya FaradisaNo ratings yet
- CV 3 Circulatory System Notes CopyrightDocument7 pagesCV 3 Circulatory System Notes Copyrightmaxwell amponsahNo ratings yet
- PART VI. Hemodynamic DisordersDocument11 pagesPART VI. Hemodynamic DisordersGideon YeboahNo ratings yet
- Blood LossDocument6 pagesBlood LossDungani AllanNo ratings yet
- Cardiovascular System: Iii. Anatomy and PhysiologyDocument7 pagesCardiovascular System: Iii. Anatomy and PhysiologyYam Bite-Size FabelloNo ratings yet
- Screenshot 2023-03-17 at 12.32.32 AMDocument1 pageScreenshot 2023-03-17 at 12.32.32 AMRenadNo ratings yet
- Anatomy EssaysDocument4 pagesAnatomy EssaysAsh HassanNo ratings yet
- Lecture 14-Oct.23 Haemodynamics of Circulation, Circulatory Mechanics, ResistanceDocument29 pagesLecture 14-Oct.23 Haemodynamics of Circulation, Circulatory Mechanics, ResistanceAparmitNo ratings yet
- Sistem SirkulasiDocument54 pagesSistem SirkulasiIhedi SacramentoNo ratings yet
- The International Journal of Engineering and Science (The IJES)Document7 pagesThe International Journal of Engineering and Science (The IJES)theijesNo ratings yet
- Renal Notes Unit 1Document66 pagesRenal Notes Unit 1Zha-Raa'iaam L SamoriNo ratings yet
- Cardiovascular Pathophysiology ExplainedDocument32 pagesCardiovascular Pathophysiology ExplainedSaif AliNo ratings yet
- Circulation. Lec On HemodynamicsDocument30 pagesCirculation. Lec On HemodynamicsMudassar RoomiNo ratings yet
- Cardiovascular Changes in CardiogenicDocument4 pagesCardiovascular Changes in CardiogenicBaso AgusofyangNo ratings yet
- Chapter 7 Regulation of Arterial Blood Pressure and MicrocirculationDocument20 pagesChapter 7 Regulation of Arterial Blood Pressure and Microcirculationaisyahasrii_No ratings yet
- HEMODINAMIKADocument42 pagesHEMODINAMIKAIndraYudhiNo ratings yet
- CASE SolutionDocument11 pagesCASE Solution037 Onpreeya LansaiNo ratings yet
- Example PVProtocol - TabletDocument22 pagesExample PVProtocol - Tablet037 Onpreeya LansaiNo ratings yet
- Process Validation Protocol of Granules For Oral AdministrationDocument6 pagesProcess Validation Protocol of Granules For Oral AdministrationRezaul haque himelNo ratings yet
- Orig 1 S 000 Med RedtDocument104 pagesOrig 1 S 000 Med Redt037 Onpreeya LansaiNo ratings yet
- SLE 2015, Compilation of Test Questions: 8 5 5 M C Q SDocument99 pagesSLE 2015, Compilation of Test Questions: 8 5 5 M C Q SAbdelreheem ElgendyNo ratings yet
- VIH-SIDA Prevension y TratamientoDocument8 pagesVIH-SIDA Prevension y TratamientoDaniel MarquezNo ratings yet
- Physiology of Micturition Reflex PDFDocument37 pagesPhysiology of Micturition Reflex PDFVanathy KarunamoorthyNo ratings yet
- Flashcards in Abo Blood GroupDocument10 pagesFlashcards in Abo Blood GroupVincent ReyesNo ratings yet
- What Happens When You Breath Niric Acid FumesDocument4 pagesWhat Happens When You Breath Niric Acid FumesAFLAC ............No ratings yet
- Maternal and Child Nursing Sample ExamDocument8 pagesMaternal and Child Nursing Sample Examteabagman0% (1)
- A Prospective Randomised Comparison of The Dynamic Hip Screw and The Gamma Locking NailDocument5 pagesA Prospective Randomised Comparison of The Dynamic Hip Screw and The Gamma Locking NailAshish UpadhyayNo ratings yet
- Maternity NotesDocument16 pagesMaternity Notesanon_551740746No ratings yet
- DELAYED SPEECH: CAUSES AND TREATMENTDocument20 pagesDELAYED SPEECH: CAUSES AND TREATMENTrizkiya novitaNo ratings yet
- Respiratory System True or False QuizDocument3 pagesRespiratory System True or False QuizDr-Atin Kumar SrivastavaNo ratings yet
- Biology Unitwise MCQ - S (MCAT)Document41 pagesBiology Unitwise MCQ - S (MCAT)M Noaman Akbar100% (1)
- History of Insulin Discovery Revolutionized Diabetes TreatmentDocument4 pagesHistory of Insulin Discovery Revolutionized Diabetes TreatmentDženan MukićNo ratings yet
- Human reproduction self-assessment questionsDocument2 pagesHuman reproduction self-assessment questionsEdyWidjajaNo ratings yet
- How Do Organisms Reproduce PDFDocument25 pagesHow Do Organisms Reproduce PDFBala VisaNo ratings yet
- Self Assessment and Review of OphthalmologyDocument221 pagesSelf Assessment and Review of OphthalmologySusobhan Paul86% (7)
- General Models in Physiology Mass Balance - Based OnDocument2 pagesGeneral Models in Physiology Mass Balance - Based OnmcwnotesNo ratings yet
- LymeDocument31 pagesLymetugurlanNo ratings yet
- Presented To: Dr. Prashant Mario D'Souza Er. Ashish Kumar Kerketta Presented By: Shivam Kumar 13BTAG118Document17 pagesPresented To: Dr. Prashant Mario D'Souza Er. Ashish Kumar Kerketta Presented By: Shivam Kumar 13BTAG118Akash SrivastavNo ratings yet
- Ans MCQ BankDocument51 pagesAns MCQ BankAmir Z Al-Mukmin75% (4)
- Limbic System NeuroscienceDocument82 pagesLimbic System Neuroscience250187155No ratings yet
- Notification of Vaccination Letter Template: Vaccines AdministeredDocument1 pageNotification of Vaccination Letter Template: Vaccines AdministeredJacob BuckNo ratings yet
- Saritha Udan Oru NalDocument10 pagesSaritha Udan Oru Nalsanti_1976No ratings yet
- Para Lab3 TapewormsDocument19 pagesPara Lab3 Tapewormsawesome avedNo ratings yet
- A Case of Subcorneal Pustular Dermatosis Successfully Treated With AcitretinDocument3 pagesA Case of Subcorneal Pustular Dermatosis Successfully Treated With Acitretindr_RMNo ratings yet
- Prenatal Development in Three Stages: Germinal, Embryonic, and FetalDocument2 pagesPrenatal Development in Three Stages: Germinal, Embryonic, and FetalMuhammad HassanNo ratings yet
- Possible PCOS Iridology Signs, Hazel Eyes - Iridology Education PDFDocument5 pagesPossible PCOS Iridology Signs, Hazel Eyes - Iridology Education PDFvidhyasree123No ratings yet
- Infecciones Virales Del SNCDocument246 pagesInfecciones Virales Del SNCpediatria hncase100% (1)
- 1 DomsDocument26 pages1 DomsDr prince prabhakarNo ratings yet
- Breaking News For Men-Effective Herbal Epididymitis TreatmentDocument10 pagesBreaking News For Men-Effective Herbal Epididymitis TreatmentwozhiainiNo ratings yet
- God's Cancer ParmacyDocument5 pagesGod's Cancer ParmacyNick BantoloNo ratings yet