You might also like
- Airway Smooth Muscle in Asthma and COPD: Biology and PharmacologyFrom EverandAirway Smooth Muscle in Asthma and COPD: Biology and PharmacologyKian Fan ChungNo ratings yet
- Stozer Et Al (2020) - Pathophysiology of Exercise Induced Muscle Damage and Its Structural, Functional, Metabolic and Clinical ConsequencesDocument34 pagesStozer Et Al (2020) - Pathophysiology of Exercise Induced Muscle Damage and Its Structural, Functional, Metabolic and Clinical ConsequencesNg Hui HwaNo ratings yet
- Stem Cells and Cell Signalling in Skeletal MyogenesisFrom EverandStem Cells and Cell Signalling in Skeletal MyogenesisNo ratings yet
- Biology of Rotator Cuff HealingDocument10 pagesBiology of Rotator Cuff HealingRaeni Dwi PutriNo ratings yet
- Physiology and BiochemistryFrom EverandPhysiology and BiochemistryGeoffrey BourneNo ratings yet
- Cardiac Preload: Physiology and Clinical ImplicationsDocument5 pagesCardiac Preload: Physiology and Clinical ImplicationsOdrig De LavalleNo ratings yet
- Book Reviews: Muscle DamageDocument3 pagesBook Reviews: Muscle DamageManisha SahNo ratings yet
- JCSM 9 631Document4 pagesJCSM 9 631Nakarit SangsirinawinNo ratings yet
- Araujo2008 PDFDocument9 pagesAraujo2008 PDFAndresNo ratings yet
- Biomechanics Upper AirwayDocument11 pagesBiomechanics Upper AirwayenNo ratings yet
- Functional Electrical Stimulation of Denervated MusclesDocument7 pagesFunctional Electrical Stimulation of Denervated MusclesMirko GrausoNo ratings yet
- Blazevich Et, Al. 2007 - Treino Concentrico Vs Treino ExcentricoDocument12 pagesBlazevich Et, Al. 2007 - Treino Concentrico Vs Treino ExcentricoFábio LanferdiniNo ratings yet
- Medi Adores Dis Fun CM LDocument7 pagesMedi Adores Dis Fun CM LdanielNo ratings yet
- Clark2008 Sacopenia DinapeniaDocument6 pagesClark2008 Sacopenia DinapeniaAna Souza LimaNo ratings yet
- Muscular Adaptations and Insulin-Like Growth FactoDocument13 pagesMuscular Adaptations and Insulin-Like Growth Factoleal thiagoNo ratings yet
- Eph 1996813329Document11 pagesEph 1996813329Bernardo SteinbergNo ratings yet
- Nieman, D. (1997) - Exercise Immunology Practical Applications. International Journal of Sports MedicineDocument10 pagesNieman, D. (1997) - Exercise Immunology Practical Applications. International Journal of Sports MedicineJeane Claudia100% (1)
- 10 1 1 1021 5805Document14 pages10 1 1 1021 5805echo_10No ratings yet
- How Do Tendons Adapt? Going Beyond Tissue Responses To Understand Positive Adaptation and Pathology Development: A Narrative ReviewDocument11 pagesHow Do Tendons Adapt? Going Beyond Tissue Responses To Understand Positive Adaptation and Pathology Development: A Narrative ReviewMário SimõesNo ratings yet
- Phys of Ex Annotated Bib FinalDocument32 pagesPhys of Ex Annotated Bib Finalapi-530529613No ratings yet
- 2014 Disfunción Diafragmática Inducida Por VentilaciónDocument8 pages2014 Disfunción Diafragmática Inducida Por VentilaciónwilsonmauriciolozanoNo ratings yet
- The Effect of Inspiratory Muscle Training On Duchenne Muscular Dystrophy: A Meta-AnalysisDocument8 pagesThe Effect of Inspiratory Muscle Training On Duchenne Muscular Dystrophy: A Meta-AnalysisTirumala RaoNo ratings yet
- Ijspt 15 579Document14 pagesIjspt 15 579Hevin GokulNo ratings yet
- 10 1152@ajpcell 00016 2016Document14 pages10 1152@ajpcell 00016 2016Muttley ZNo ratings yet
- A Comparative Study To Find Out The Effects of Capsular Stretching Over Muscle Energy Technique in The Management of Frozen ShoulderDocument50 pagesA Comparative Study To Find Out The Effects of Capsular Stretching Over Muscle Energy Technique in The Management of Frozen ShoulderjojiNo ratings yet
- A Review of The Role of Mechanical Forces in Cutaneous Wound HealingDocument9 pagesA Review of The Role of Mechanical Forces in Cutaneous Wound HealingAna MelloNo ratings yet
- European Journal of PharmacologyDocument9 pagesEuropean Journal of PharmacologyMarcelo Pires de OliveiraNo ratings yet
- Saladin: Anatomy & Physiology: The Unity of Form and Function, Third Edition Front Matter Clinical EmphasisDocument3 pagesSaladin: Anatomy & Physiology: The Unity of Form and Function, Third Edition Front Matter Clinical EmphasisJelena PopovićNo ratings yet
- Stress, Aging, and Resilience: Can Accrued Wear and Tear Be Slowed?Document11 pagesStress, Aging, and Resilience: Can Accrued Wear and Tear Be Slowed?bastian1976No ratings yet
- Biomechanics of Nerve 211080006 Rehab AymanDocument9 pagesBiomechanics of Nerve 211080006 Rehab Aymanrehab aymanNo ratings yet
- Molecular Pain: Short-Term Synaptic Plasticity in The Nociceptive Thalamic-Anterior Cingulate PathwayDocument20 pagesMolecular Pain: Short-Term Synaptic Plasticity in The Nociceptive Thalamic-Anterior Cingulate PathwayYixin TongNo ratings yet
- PTJ 0357Document19 pagesPTJ 0357Jose JuniorNo ratings yet
- Enlargement Resulting From Surgical Intervention Chronic Static Magnetic Field Exposure Alters MicrovesselDocument9 pagesEnlargement Resulting From Surgical Intervention Chronic Static Magnetic Field Exposure Alters MicrovesselSanders Perfum CosmetNo ratings yet
- Skeletal Muscle Damage and RecoveryDocument11 pagesSkeletal Muscle Damage and Recoveryrolinga123No ratings yet
- TMP 429 FDocument17 pagesTMP 429 FFrontiersNo ratings yet
- J MusculoskPainDocument4 pagesJ MusculoskPainBruno OliveiraNo ratings yet
- Fascia Can Contract in Smooth Muscle Like Manner PDFDocument7 pagesFascia Can Contract in Smooth Muscle Like Manner PDFVeronica Petit LamedaNo ratings yet
- Exercise-Induced Central Fatigue: A Review of The Literature With Implications For Dance Science ResearchDocument10 pagesExercise-Induced Central Fatigue: A Review of The Literature With Implications For Dance Science ResearchShyamali Kaushalya FernandoNo ratings yet
- Subscapularis Sy DonatelliDocument12 pagesSubscapularis Sy DonatelliDusan OrescaninNo ratings yet
- A Systematic Review of The Effectiveness of Eccentric Strength Training in The Prevention of Hamstring Muscle Strains in Otherwise Healthy IndividualsDocument15 pagesA Systematic Review of The Effectiveness of Eccentric Strength Training in The Prevention of Hamstring Muscle Strains in Otherwise Healthy IndividualsTiago RochaNo ratings yet
- Mechanical Loads - Bone ResponseDocument19 pagesMechanical Loads - Bone ResponselllllAdelalllllNo ratings yet
- 2 VentriclesDocument45 pages2 Ventricles15xicoNo ratings yet
- Biofeedback Therapy in Stroke Rehabilitation: Review: Morton Glanz Sidney Klawansky Thomas ChalmersDocument7 pagesBiofeedback Therapy in Stroke Rehabilitation: Review: Morton Glanz Sidney Klawansky Thomas Chalmersnandhini raguNo ratings yet
- Mechanisms of Blood Flow Restriction: The New Testament: YmposiumDocument8 pagesMechanisms of Blood Flow Restriction: The New Testament: YmposiumRosaneLacerdaNo ratings yet
- Action of The Diaphragm On The Rib Cage2016Document10 pagesAction of The Diaphragm On The Rib Cage2016Adynna TévinaNo ratings yet
- Effects of Spasticity On Motor Recovery After A StrokeDocument10 pagesEffects of Spasticity On Motor Recovery After A StrokeDaniel SanchezNo ratings yet
- Voice Rest Versus Exercise: A Review of The Literature: Madison, WisconsinDocument9 pagesVoice Rest Versus Exercise: A Review of The Literature: Madison, WisconsinJaime Crisosto AlarcónNo ratings yet
- Visceral Smooth MuscleDocument3 pagesVisceral Smooth MusclehardianNo ratings yet
- Artigo FesDocument19 pagesArtigo FesThiago Penna ChavesNo ratings yet
- Effects of Mechanical Factors On The Fracture Healing ProcessDocument16 pagesEffects of Mechanical Factors On The Fracture Healing ProcessBastion ReinhartNo ratings yet
- Dosen Pengampu Mata Kuliah Fisiotherapy: Dr. Konni Kurniasih, M.Kes., AIFODocument7 pagesDosen Pengampu Mata Kuliah Fisiotherapy: Dr. Konni Kurniasih, M.Kes., AIFOilham POYNo ratings yet
- ClinBiomech 16 87-101 2001Document15 pagesClinBiomech 16 87-101 2001Paulo GomesNo ratings yet
- Proposal s3Document11 pagesProposal s3RifqiNo ratings yet
- Respiratory Diseases and Muscle Dysfunction: ReviewDocument17 pagesRespiratory Diseases and Muscle Dysfunction: ReviewMatias Emilio BirknerNo ratings yet
- Fitzpatrick 2016Document13 pagesFitzpatrick 2016smansa123No ratings yet
- Jan HoffDocument7 pagesJan Hofflsales1986oliNo ratings yet
- Evaluacion de Los Musculos Respiratorios en La Practica ClinicaDocument9 pagesEvaluacion de Los Musculos Respiratorios en La Practica ClinicaDaniela CalleNo ratings yet
- Mohamed Abdalla 211080013Document11 pagesMohamed Abdalla 211080013rehab aymanNo ratings yet
- Deficit SensoriomotorDocument7 pagesDeficit SensoriomotorAnonymous FWF4P38wHNo ratings yet
- Lacroix 2002Document9 pagesLacroix 2002Bastion ReinhartNo ratings yet
- Li 2019Document28 pagesLi 2019Jocilene Dantas Torres NascimentoNo ratings yet
- Shay 2011Document5 pagesShay 2011Jocilene Dantas Torres NascimentoNo ratings yet
- Nair 2016Document1 pageNair 2016Jocilene Dantas Torres NascimentoNo ratings yet
- Koch 2016Document2 pagesKoch 2016Jocilene Dantas Torres NascimentoNo ratings yet
- Cole 2016Document1 pageCole 2016Jocilene Dantas Torres NascimentoNo ratings yet
- Mattei 1985Document5 pagesMattei 1985Jocilene Dantas Torres NascimentoNo ratings yet
- Gruca 1995Document12 pagesGruca 1995Jocilene Dantas Torres NascimentoNo ratings yet
- False-Positive Alkaloid ReactionsDocument7 pagesFalse-Positive Alkaloid ReactionsJocilene Dantas Torres NascimentoNo ratings yet
- Zhou 2018Document21 pagesZhou 2018Jocilene Dantas Torres NascimentoNo ratings yet
- Physiological and Psychological FactorsDocument12 pagesPhysiological and Psychological FactorsJocilene Dantas Torres NascimentoNo ratings yet
- Theology For YouthDocument10 pagesTheology For YouthJocilene Dantas Torres NascimentoNo ratings yet
- Quartz Disrupts Iron HomeostasisDocument11 pagesQuartz Disrupts Iron HomeostasisJocilene Dantas Torres NascimentoNo ratings yet
- Glycyrrhizic AcidDocument8 pagesGlycyrrhizic AcidJocilene Dantas Torres NascimentoNo ratings yet
- Ballash 2006Document21 pagesBallash 2006Jocilene Dantas Torres NascimentoNo ratings yet
- Thompson 1997Document1 pageThompson 1997Jocilene Dantas Torres NascimentoNo ratings yet
- Leone1998-Cervicogenic HeadacheDocument5 pagesLeone1998-Cervicogenic HeadacheJocilene Dantas Torres NascimentoNo ratings yet
- Mcneice 2020Document6 pagesMcneice 2020Jocilene Dantas Torres NascimentoNo ratings yet
- Journal of Membrane Science: 2 Qi Zhong, Guogui Shi, Qing Sun, Peng Mu, Jian LiDocument11 pagesJournal of Membrane Science: 2 Qi Zhong, Guogui Shi, Qing Sun, Peng Mu, Jian LiJocilene Dantas Torres NascimentoNo ratings yet
- Review Article Ancient China Reconsidered: Magnus FiskesjöDocument3 pagesReview Article Ancient China Reconsidered: Magnus FiskesjöJocilene Dantas Torres NascimentoNo ratings yet
- Suda 2018Document12 pagesSuda 2018Jocilene Dantas Torres NascimentoNo ratings yet
- Diagnosis and Management of Acute Ischaemic StrokDocument13 pagesDiagnosis and Management of Acute Ischaemic StroksamuelNo ratings yet
- Seong 2015Document7 pagesSeong 2015Jocilene Dantas Torres NascimentoNo ratings yet
- Physico-Chemical Properties of Kenyan Made Calcined Clay - Limestone Cement (LC3)Document13 pagesPhysico-Chemical Properties of Kenyan Made Calcined Clay - Limestone Cement (LC3)Angel Samir Diaz PedrazaNo ratings yet
- Iso 1871 2009Document9 pagesIso 1871 2009Jocilene Dantas Torres NascimentoNo ratings yet
- Catálogo Gerber Cap-LabDocument83 pagesCatálogo Gerber Cap-LabJocilene Dantas Torres NascimentoNo ratings yet
- Khan 2020Document11 pagesKhan 2020Jocilene Dantas Torres NascimentoNo ratings yet
- Analytical Notice Lovi 04 16 GBDocument2 pagesAnalytical Notice Lovi 04 16 GBJocilene Dantas Torres NascimentoNo ratings yet
- Cenizas Leche PDFDocument1 pageCenizas Leche PDFerika vanesa higuera posadaNo ratings yet
- Live DeadDocument8 pagesLive DeadAshish KushwahaNo ratings yet
- Trauma and The Brain 1Document14 pagesTrauma and The Brain 1Syed Shams Haider MashhadiNo ratings yet
- PPE Physical Exam Form Single PageDocument1 pagePPE Physical Exam Form Single PageyumminNo ratings yet
- Chapter 2 BotanyDocument6 pagesChapter 2 BotanyLebanan Aprille MarieNo ratings yet
- Digestive SystemDocument17 pagesDigestive SystemTricia GorozaNo ratings yet
- Nursing Review Exam Questions FUNDADocument10 pagesNursing Review Exam Questions FUNDAAlibasher MacalnasNo ratings yet
- Bantuan Hidup DasarDocument6 pagesBantuan Hidup DasarRamadhanaNo ratings yet
- ReninDocument4 pagesReninSarah AndrianiNo ratings yet
- Intro Asanas BookletDocument5 pagesIntro Asanas BookletthegreatmanNo ratings yet
- Equ211-01 Siemens Dimension Fast FactsDocument26 pagesEqu211-01 Siemens Dimension Fast Factsmagendi indra muktiNo ratings yet
- Sucrose Gradient Separation ProtocolDocument16 pagesSucrose Gradient Separation ProtocolabatabrahamNo ratings yet
- Abdominal Wall: Muh. Iqbal Basri Anatomy Department Hasanuddin UniversityDocument33 pagesAbdominal Wall: Muh. Iqbal Basri Anatomy Department Hasanuddin UniversityDavid ChristianNo ratings yet
- Nasal SurgeryDocument3 pagesNasal SurgeryhonnyzeNo ratings yet
- Doppler Artifacts and PitfallsDocument31 pagesDoppler Artifacts and PitfallsButton Davidson100% (1)
- Cytokines, Cytokine Receptors and Chemokines: Sept. 11, 2014Document101 pagesCytokines, Cytokine Receptors and Chemokines: Sept. 11, 2014ANJU0709No ratings yet
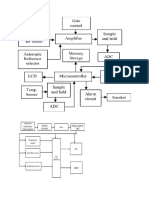
- Block Diagram PT MonitorDocument7 pagesBlock Diagram PT MonitorAbabil Academy KannanNo ratings yet
- Case Study HMoleDocument7 pagesCase Study HMolekarl_poorNo ratings yet
- Chapter 2. Acute and Chronic InflammationDocument6 pagesChapter 2. Acute and Chronic InflammationWaseem Khan AfridiNo ratings yet
- Chapter 4 - Explant Preparation - 2013 - Plant Tissue Culture PDFDocument7 pagesChapter 4 - Explant Preparation - 2013 - Plant Tissue Culture PDFLuis BermudezNo ratings yet
- 11 класс 4 модульDocument4 pages11 класс 4 модульYuriNo ratings yet
- Unit 2Document8 pagesUnit 2Mason BruceNo ratings yet
- Chapter 15 PLANT GROWTH AND DEVELOPMENT Exercises PDFDocument33 pagesChapter 15 PLANT GROWTH AND DEVELOPMENT Exercises PDFHoney Giri goswami100% (1)
- Blood Everywhere: A Case Study in BloodDocument2 pagesBlood Everywhere: A Case Study in BloodAbdullahNo ratings yet
- Assessing Thorax LungsDocument20 pagesAssessing Thorax Lungskyla boncacasNo ratings yet
- Foundations of Fitness ProgrammingDocument35 pagesFoundations of Fitness ProgrammingPaul Rose75% (4)
- IntroductionDocument2 pagesIntroductionHarvey Gian JavierNo ratings yet
- Pharmacology-II NIRALIDocument244 pagesPharmacology-II NIRALIRock Patel93% (29)
- CBC ReportsDocument2 pagesCBC ReportsIbrar Hussain100% (1)
- Breathing ExercisesDocument16 pagesBreathing ExercisesSubramanya Seshagiri100% (1)
- C - Quick Access Table - Combining Forms To Definitions 1Document19 pagesC - Quick Access Table - Combining Forms To Definitions 1neuronurseNo ratings yet
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (29)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (2)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (404)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (81)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (170)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)From EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)No ratings yet
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (6)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeRating: 2 out of 5 stars2/5 (1)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (4)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- Summary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisFrom EverandSummary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisRating: 5 out of 5 stars5/5 (8)
- To Explain the World: The Discovery of Modern ScienceFrom EverandTo Explain the World: The Discovery of Modern ScienceRating: 3.5 out of 5 stars3.5/5 (51)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- The Marshmallow Test: Mastering Self-ControlFrom EverandThe Marshmallow Test: Mastering Self-ControlRating: 4.5 out of 5 stars4.5/5 (58)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (45)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (253)