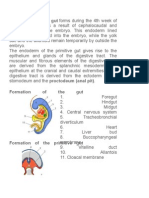
You might also like
- As Epidermal Stem Cells Age They Do Not Substantially Change Their CharacteristicsDocument9 pagesAs Epidermal Stem Cells Age They Do Not Substantially Change Their CharacteristicsMichał PikułaNo ratings yet
- Ectopic Thyroid: Etiology, Pathology and Management: ReviewDocument9 pagesEctopic Thyroid: Etiology, Pathology and Management: ReviewIonuț RusNo ratings yet
- The Chicken As A Model For Embryonic Development: M.G. Davey C. TickleDocument9 pagesThe Chicken As A Model For Embryonic Development: M.G. Davey C. TickleUtari Septiana DewiNo ratings yet
- Fate MappingDocument45 pagesFate MappingDale Marie RenomeronNo ratings yet
- Germline Regulatory Element of Oct-4 Specific For The Totipotent Cycle ofDocument14 pagesGermline Regulatory Element of Oct-4 Specific For The Totipotent Cycle ofMariane Gabriela Cesar Ribeiro FerreiraNo ratings yet
- Reproductive Biology and Endocrinology: Oogenesis in Cultures Derived From Adult Human OvariesDocument13 pagesReproductive Biology and Endocrinology: Oogenesis in Cultures Derived From Adult Human Ovaries05-OB-HU-DINA MERCEDES CAMPOSANO SALAZARNo ratings yet
- Jurnal ObstetriDocument9 pagesJurnal ObstetriGhaly KhaidarNo ratings yet
- VishuDocument4 pagesVishuVishuNo ratings yet
- New Insights Into Human Early Embryo Development: A Particular Theoretical StudyDocument33 pagesNew Insights Into Human Early Embryo Development: A Particular Theoretical StudyIván DelgadoNo ratings yet
- Lecture Guide: Introduction To EmbryologyDocument23 pagesLecture Guide: Introduction To EmbryologyAldrin Hardy PabloNo ratings yet
- Human EmbriologyDocument67 pagesHuman EmbriologyGalang Perdamaian0% (1)
- What Is An Embryo (Inglés) Autor Anna A. KiesslingDocument42 pagesWhat Is An Embryo (Inglés) Autor Anna A. Kiesslingredactor 1No ratings yet
- Embryo Technology: Implications For Fertility in Cattle: T. Greve & H. CallesenDocument8 pagesEmbryo Technology: Implications For Fertility in Cattle: T. Greve & H. CallesenmuniveleNo ratings yet
- Embryology of The Hind GutDocument10 pagesEmbryology of The Hind Gutproject-247758No ratings yet
- Biology: Find and Fuse: Unsolved Mysteries in Sperm - Egg RecognitionDocument11 pagesBiology: Find and Fuse: Unsolved Mysteries in Sperm - Egg RecognitionFlavio NavarreteNo ratings yet
- Sonographic Diagnosis of Placental Teratoma: Case ReportDocument4 pagesSonographic Diagnosis of Placental Teratoma: Case ReportDen JCNo ratings yet
- Safe Female CircumcisionDocument51 pagesSafe Female Circumcisiond7rqqrk+vaii68100% (1)
- Ti PDFDocument9 pagesTi PDFAbanx Wiko ExperiencedNo ratings yet
- Structural Changes in Endometrial Basal Glands During MenstruationDocument11 pagesStructural Changes in Endometrial Basal Glands During MenstruationWahab RopekNo ratings yet
- Thymic Microenvironments, 3-D Versus 2-D?: 64 Article No. Smim.1998.0158, Available Online at HTTPDocument8 pagesThymic Microenvironments, 3-D Versus 2-D?: 64 Article No. Smim.1998.0158, Available Online at HTTPpedroNo ratings yet
- Chapter 33 - Descriptive EmbryologyDocument12 pagesChapter 33 - Descriptive Embryologyngquvu67% (15)
- Gal Et Al HaydarDocument12 pagesGal Et Al HaydarfloodkNo ratings yet
- 403 2009 Article 989-1 PDFDocument9 pages403 2009 Article 989-1 PDFUtari Septiana DewiNo ratings yet
- Ovariancystsintheguineapig (Caviaporcellus) : Andrew D. BeanDocument20 pagesOvariancystsintheguineapig (Caviaporcellus) : Andrew D. BeanALEX MOREANO ACOSTUPANo ratings yet
- Cell Culture HandbookDocument30 pagesCell Culture HandbookRamasamy VelmuruganNo ratings yet
- Unlock SGCh22Document8 pagesUnlock SGCh22SilaxNo ratings yet
- Review of Prostate Anatomy and Embryology and The Etiology of Benign Prostatic Hyperplasia PDFDocument10 pagesReview of Prostate Anatomy and Embryology and The Etiology of Benign Prostatic Hyperplasia PDFAlejandro GuerreroNo ratings yet
- 45 HypospadiaDocument50 pages45 HypospadiaMarcella SaputraNo ratings yet
- Sofie R Krisnadi, Dr. SpogDocument82 pagesSofie R Krisnadi, Dr. SpogYogi OktiandiNo ratings yet
- Yolk Protein Uptake in The Oocyte of The Mosquito: Aedes Aegypti. LDocument20 pagesYolk Protein Uptake in The Oocyte of The Mosquito: Aedes Aegypti. LRogério HitsugayaNo ratings yet
- Cell RevolutionDocument13 pagesCell RevolutionDream KyleNo ratings yet
- Molecular Mechanism of ImplantationDocument5 pagesMolecular Mechanism of Implantationarunnene2No ratings yet
- Biological Basis For Human Capacitation: Christopher de JongeDocument10 pagesBiological Basis For Human Capacitation: Christopher de JongeVicNo ratings yet
- Biological Basis For Human Capacitation: Christopher de JongeDocument10 pagesBiological Basis For Human Capacitation: Christopher de JongeLara RamundoNo ratings yet
- J Bbagrm 2009 02 002Document7 pagesJ Bbagrm 2009 02 002Raymundo Lopez NNo ratings yet
- Synthesis Oxytocin Amnion, Chorion, Influence The Timing of HumanDocument8 pagesSynthesis Oxytocin Amnion, Chorion, Influence The Timing of HumanElissa IsdasariNo ratings yet
- Ajpendo 00705 2006Document8 pagesAjpendo 00705 2006Fujiko SaavedraNo ratings yet
- Great Sperm RaceDocument19 pagesGreat Sperm RacefrjesNo ratings yet
- The Transient Cortical Zone in The Adrenal Gland: The Mystery of The Adrenal X-ZoneDocument13 pagesThe Transient Cortical Zone in The Adrenal Gland: The Mystery of The Adrenal X-ZoneCandle Jo'inNo ratings yet
- Acta Obstet Gynecol Scand - 2011 - LIND - Collagens in The Human Ovary and Their Changes in The Perifollicular StromaDocument9 pagesActa Obstet Gynecol Scand - 2011 - LIND - Collagens in The Human Ovary and Their Changes in The Perifollicular StromaalnyhilwanyNo ratings yet
- Farin 2009Document25 pagesFarin 2009Laura Duarte RojasNo ratings yet
- The Placenta and Fetal MembranesDocument69 pagesThe Placenta and Fetal MembranesAnu ChauhanNo ratings yet
- Inv 14Document8 pagesInv 14mrojasf.1996No ratings yet
- Derivation of Totipotent-Like Stem Cells With Blastocyst-Like Structure Forming PotentialDocument17 pagesDerivation of Totipotent-Like Stem Cells With Blastocyst-Like Structure Forming PotentialJosué Cristhian Del Valle HornaNo ratings yet
- HAMILDocument82 pagesHAMILNandya CarolineNo ratings yet
- Building The Mammalian Testis: Origins, Differentiation, and Assembly of The Component Cell PopulationsDocument19 pagesBuilding The Mammalian Testis: Origins, Differentiation, and Assembly of The Component Cell Populationssendy ali baehaqiNo ratings yet
- Version On-Line ISSN 0717-9502: International Journal of MorphologyDocument8 pagesVersion On-Line ISSN 0717-9502: International Journal of MorphologyMaría AlejandraNo ratings yet
- Placenta: J.L. James, S. Srinivasan, M. Alexander, L.W. ChamleyDocument8 pagesPlacenta: J.L. James, S. Srinivasan, M. Alexander, L.W. ChamleyfujimeisterNo ratings yet
- Information Networks in The Mammary Gland: Lothar Hennighausen and Gertraud W. RobinsonDocument11 pagesInformation Networks in The Mammary Gland: Lothar Hennighausen and Gertraud W. RobinsonSueños De LunaNo ratings yet
- Jelkd Jeo QWNDocument25 pagesJelkd Jeo QWNcd_armand1885No ratings yet
- Stem Cell Technology: (Module in Cell Molecular Biology)Document23 pagesStem Cell Technology: (Module in Cell Molecular Biology)Anonymous H2L7lwBs3No ratings yet
- Stem Cell Technology: (Module in Cell Molecular Biology)Document8 pagesStem Cell Technology: (Module in Cell Molecular Biology)Anonymous H2L7lwBs3No ratings yet
- Nervous Control of Ovulation and Ejaculation in Helix AspersaDocument7 pagesNervous Control of Ovulation and Ejaculation in Helix AspersaCarlos MeirellesNo ratings yet
- ISSN 1679-9216 (Online)Document19 pagesISSN 1679-9216 (Online)Rachel HayonNo ratings yet
- First Page PDFDocument1 pageFirst Page PDFAdelaide Lucien CarrissaNo ratings yet
- 2002 - The Role of Colesterol Efflux in Regulating The Fertilization Potential of Mammalian SpermatozoaDocument7 pages2002 - The Role of Colesterol Efflux in Regulating The Fertilization Potential of Mammalian Spermatozoadavid torresNo ratings yet
- 1 s2.0 S0143400411005339 Main PDFDocument8 pages1 s2.0 S0143400411005339 Main PDFMarco Antonio AspronNo ratings yet
- 180 Female and Male Gametogenesis588Document20 pages180 Female and Male Gametogenesis588Hari DonNo ratings yet
- Determinants of Postnatal Spleen Tissue Regeneration and OrganogenesisDocument4 pagesDeterminants of Postnatal Spleen Tissue Regeneration and OrganogenesisDaniel Daza BarriosNo ratings yet
- Brief Readings SWEDENBORG's PSYCHOLOGY Howard Davis Spoerl Swedenborg Foundation 1937Document16 pagesBrief Readings SWEDENBORG's PSYCHOLOGY Howard Davis Spoerl Swedenborg Foundation 1937francis batt100% (1)
- Adv Acu Tech 1 - The Second Metacarpal Bone TherapyDocument24 pagesAdv Acu Tech 1 - The Second Metacarpal Bone Therapysuperser123465100% (2)
- Heart Failure Clinical Records DatasetDocument14 pagesHeart Failure Clinical Records Datasetpanca saputraNo ratings yet
- Data Alkes 2020 TerbaruDocument23 pagesData Alkes 2020 TerbaruRoyliNo ratings yet
- The Workings of The Brain Powers of The MindDocument17 pagesThe Workings of The Brain Powers of The MindLovely ramirezNo ratings yet
- Raddaily Ankle MRI ApproachDocument10 pagesRaddaily Ankle MRI ApproachSandeep VodelaNo ratings yet
- Testicular CancerDocument5 pagesTesticular CancerypintohNo ratings yet
- Think of Yourself As A KingDocument7 pagesThink of Yourself As A Kingiyayi2006No ratings yet
- Vedda+Blood+Sugar+Remedy+PDF+ +Book+Recipes+Free+Download+Michael+DempseyDocument20 pagesVedda+Blood+Sugar+Remedy+PDF+ +Book+Recipes+Free+Download+Michael+DempseyBianca SmitheIncas AllenNo ratings yet
- Final Compre Exam Key AnswersDocument14 pagesFinal Compre Exam Key Answersmj CanilangNo ratings yet
- Kreisberg-Zakarin Et Al IPNS Antonie Van Leeuwenhoek 1999Document8 pagesKreisberg-Zakarin Et Al IPNS Antonie Van Leeuwenhoek 1999ElNo ratings yet
- Bp503t Pcol Unit-VDocument46 pagesBp503t Pcol Unit-VAakkkNo ratings yet
- Rula SheetDocument2 pagesRula SheetCarl Christian CabelloNo ratings yet
- Knight, Angela - The VampdomDocument8 pagesKnight, Angela - The VampdomLaviniaNo ratings yet
- Class XI Practicals Term2Document5 pagesClass XI Practicals Term2Prince KNo ratings yet
- Human Anatomy 4th Edition McKinley Test Bank 1Document35 pagesHuman Anatomy 4th Edition McKinley Test Bank 1donnawugnwsjrzcxt100% (27)
- 2-Epithelium MCQ Asnan Ainshams DR - Zahra 2020Document9 pages2-Epithelium MCQ Asnan Ainshams DR - Zahra 2020Shay Os100% (1)
- 6.1 Digestion BNDocument24 pages6.1 Digestion BNHana ElattarNo ratings yet
- Respiration Lecture NotesDocument7 pagesRespiration Lecture Notesamrutha mohandasNo ratings yet
- Steroids: Understanding Causes, Effects, and InformationDocument9 pagesSteroids: Understanding Causes, Effects, and InformationChris weberNo ratings yet
- CPG THDocument335 pagesCPG THAhmad Syahmi YZNo ratings yet
- Brooke Cybel TanateDocument5 pagesBrooke Cybel TanateEbenezer Manguerra Diala Gube Jr.No ratings yet
- Anaesthetic Implications in A Case of Indian Patient of Xeroderma PigmentosumDocument5 pagesAnaesthetic Implications in A Case of Indian Patient of Xeroderma PigmentosumSwapnil AswarNo ratings yet
- Sorenson Atlas of Human Histology Chapters 1 and 14Document56 pagesSorenson Atlas of Human Histology Chapters 1 and 14Nick john CaminadeNo ratings yet
- Bio Quiz BeeDocument2 pagesBio Quiz BeeRuchi Briam James LagitnayNo ratings yet
- Co2 PDFDocument4 pagesCo2 PDFSITI SARAH BTE NOOR MEAHNo ratings yet
- Patterns of Electron Flow Through Light Reaction EventsDocument25 pagesPatterns of Electron Flow Through Light Reaction EventsMar'J0% (1)
- Acute Miocard InfarkDocument32 pagesAcute Miocard InfarkhansNo ratings yet
- Voicedisordersinthe Elderly: Karen M. Kost,, Robert T. SataloffDocument13 pagesVoicedisordersinthe Elderly: Karen M. Kost,, Robert T. SataloffFernanda MerichesNo ratings yet
- Lea Joy Jugo Exercise 23Document6 pagesLea Joy Jugo Exercise 23Joan Aclo SerencioNo ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (516)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4 out of 5 stars4/5 (5)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)