You might also like
- Biopharmaceutical 1Document78 pagesBiopharmaceutical 1Pharmacy Admission ExpertNo ratings yet
- Modification and Degradation of ProteinDocument6 pagesModification and Degradation of ProteinMuhammad Usman TahirNo ratings yet
- Human Peptide - Insulin-1Document35 pagesHuman Peptide - Insulin-1Jun KooNo ratings yet
- Induced Expression Strategies and ProtocolsDocument61 pagesInduced Expression Strategies and ProtocolsAbhi DeshmukhNo ratings yet
- In Vitro TranslationDocument19 pagesIn Vitro TranslationlaraibNo ratings yet
- 10 Recombinant ProteinsDocument28 pages10 Recombinant Proteinsanshuhere0No ratings yet
- A3.Proteins 1Document46 pagesA3.Proteins 1ÇağlaNo ratings yet
- Biochem Seminar FinalDocument42 pagesBiochem Seminar FinalrukanringshawlNo ratings yet
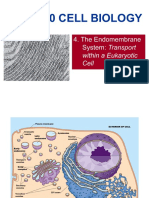
- BIOL2120 4 Endomembrane SystemDocument47 pagesBIOL2120 4 Endomembrane SystemHui Ka HoNo ratings yet
- EnzymesDocument23 pagesEnzymesBalram NeupaneNo ratings yet
- BEU11403 Biomaterials - Lecture 2 PDFDocument40 pagesBEU11403 Biomaterials - Lecture 2 PDFKin HamzahNo ratings yet
- Vipul Guddhe 7.20 BiochemDocument11 pagesVipul Guddhe 7.20 BiochemVIPul GuddheNo ratings yet
- HW5 EnzymesDocument2 pagesHW5 EnzymesFATIMA AIRA LEGASPINo ratings yet
- Produksi Protein RekombinanDocument40 pagesProduksi Protein RekombinanRaka1793No ratings yet
- Classification of ProtiensDocument4 pagesClassification of ProtiensSarah SuataronNo ratings yet
- Handout Biokimia LanjutDocument111 pagesHandout Biokimia Lanjuttheresia nababanNo ratings yet
- Cells Factories: Choosing The Right Host For Biomass ProductionDocument6 pagesCells Factories: Choosing The Right Host For Biomass ProductionalessandraNo ratings yet
- Protein Trafficking and TargettingDocument13 pagesProtein Trafficking and TargettingsoundaryaNo ratings yet
- Chapter Protein Folding and MisfoldingDocument17 pagesChapter Protein Folding and MisfoldingMazhar FarNo ratings yet
- Metabolic EngineeringDocument24 pagesMetabolic EngineeringHitaxi Dave0% (1)
- 16 HF 114 Introduction To MetabolismDocument26 pages16 HF 114 Introduction To Metabolismqueenmasa191No ratings yet
- Evelyn Gabriela Gonzalez SimbañaDocument6 pagesEvelyn Gabriela Gonzalez SimbañaMOnzerratheManzzanitaSixxNo ratings yet
- Production of Recombinant Therapeutic ProteinsDocument28 pagesProduction of Recombinant Therapeutic ProteinstasniyanNo ratings yet
- Recombinant Protein Production in E.coliDocument23 pagesRecombinant Protein Production in E.coliSanjay KumarNo ratings yet
- 1 BiopharmaceuticalDocument97 pages1 BiopharmaceuticalAnjali SaigalNo ratings yet
- ACCTDocument12 pagesACCTmAx GamiNgNo ratings yet
- Biopharmaceuticals Chapter 2-2023Document191 pagesBiopharmaceuticals Chapter 2-2023Phan Thị Minh TrúcNo ratings yet
- Erythrosomes: Pinank V. Pandya Babaria Institute of Pharmacy, VadodaraDocument21 pagesErythrosomes: Pinank V. Pandya Babaria Institute of Pharmacy, VadodaraSagar Patel0% (2)
- Smooth Endoplasmic ReticulumDocument7 pagesSmooth Endoplasmic Reticulummishiii1006343No ratings yet
- Biochem NursingDocument144 pagesBiochem Nursingabukaritoyibu100No ratings yet
- Ich Q5 Guidelines: Stability Testing of Biological/Biotechnological ProductsDocument38 pagesIch Q5 Guidelines: Stability Testing of Biological/Biotechnological ProductsSai BinduNo ratings yet
- Ich q5 GuidelinesDocument38 pagesIch q5 GuidelinesSai BinduNo ratings yet
- Post Translational Modification of Protein: Krishnakumar M S M.SC BiochemistryDocument17 pagesPost Translational Modification of Protein: Krishnakumar M S M.SC BiochemistryKrishna KumarNo ratings yet
- Endomembrane SystemDocument67 pagesEndomembrane SystemjhanvisNo ratings yet
- 24 Cellular SignalingDocument37 pages24 Cellular SignalingGian BanaresNo ratings yet
- Proteins MetabolismDocument27 pagesProteins MetabolismFouzia GillNo ratings yet
- Molecular PharmingDocument27 pagesMolecular PharmingBalakrishnan NatarajanNo ratings yet
- Biochemistry - Protein and LipidsDocument31 pagesBiochemistry - Protein and LipidsGiorgi TamazashviliNo ratings yet
- Anabolism - Microbial MetabolismDocument28 pagesAnabolism - Microbial MetabolismDni AwatifNo ratings yet
- Modification of Proteins: Bimlesh MannDocument30 pagesModification of Proteins: Bimlesh MannAmit Kr GodaraNo ratings yet
- Hormones Lecture 3 HandoutDocument17 pagesHormones Lecture 3 HandoutFaisal_Khatib_juNo ratings yet
- Protein Degradation, Molecular PathwaysDocument10 pagesProtein Degradation, Molecular PathwaysShubhAm PalNo ratings yet
- Seminar On Protein and Peptides Drug DeliveryDocument37 pagesSeminar On Protein and Peptides Drug DeliverykeyurNo ratings yet
- Structures and Functions of ProteinDocument54 pagesStructures and Functions of ProteinRollyRiksantoNo ratings yet
- Digestion Absorpton of ProteinsDocument37 pagesDigestion Absorpton of Proteinsmohammed aliNo ratings yet
- Biochemistry & Genetics: II SHS 109: Resource Person: DR Tanveer Akbar Reference TextDocument60 pagesBiochemistry & Genetics: II SHS 109: Resource Person: DR Tanveer Akbar Reference TexttNo ratings yet
- 02 - Classification and Functions of Simple and Complex ProteinsDocument34 pages02 - Classification and Functions of Simple and Complex ProteinselizaviraniNo ratings yet
- Cytosol 1Document20 pagesCytosol 1Swastu Nurul AzizahNo ratings yet
- Denaturation and Renaturation of Ribonuclease - Unit4 A - Lecture - Unit 4Document18 pagesDenaturation and Renaturation of Ribonuclease - Unit4 A - Lecture - Unit 4raghuvanshiparnika4No ratings yet
- Bio 310Document39 pagesBio 310flipside1o1No ratings yet
- Post-Translational Modification: Dr. Md. Imtaiyaz HassanDocument61 pagesPost-Translational Modification: Dr. Md. Imtaiyaz Hassanbiotechnology2007100% (1)
- CellDocument37 pagesCellKrish NatbhanjanNo ratings yet
- Membrane ProteinsDocument47 pagesMembrane ProteinsStanley ChikoveNo ratings yet
- Fermentation Process Events Affecting The Biopharmaceutical Quality1Document10 pagesFermentation Process Events Affecting The Biopharmaceutical Quality1Haroon KundiNo ratings yet
- TT SHPTDocument35 pagesTT SHPTfeliknguyenNo ratings yet
- Cell PhysiologyDocument45 pagesCell PhysiologyHimani JhaNo ratings yet
- Regulation ReportDocument14 pagesRegulation Reportjeffersonmanalo787No ratings yet
- Recombinant Protein ProductionDocument23 pagesRecombinant Protein ProductionAnamitra SenNo ratings yet
- NO.11 1. Golgi ApparatusDocument25 pagesNO.11 1. Golgi ApparatusEkaterina SurguladzeNo ratings yet
- A-level Biology Revision: Cheeky Revision ShortcutsFrom EverandA-level Biology Revision: Cheeky Revision ShortcutsRating: 5 out of 5 stars5/5 (5)
- T4. Monoclonal AntibodiesDocument3 pagesT4. Monoclonal Antibodiesjaviergr1122000hermanodediegoNo ratings yet
- Inflammation 3Document12 pagesInflammation 3Bhavya KakkarNo ratings yet
- The Cell in Its Environment SummaryDocument1 pageThe Cell in Its Environment Summaryapi-292648627100% (1)
- Cancer Cell Biology SyllabusDocument2 pagesCancer Cell Biology SyllabusJose-Rico GonzalesNo ratings yet
- Ch. 34 Eoc ReviewDocument12 pagesCh. 34 Eoc ReviewQussayNo ratings yet
- ToeflDocument7 pagesToeflNadia TussolehaNo ratings yet
- Trial Spm-Bio k2 SkemaDocument14 pagesTrial Spm-Bio k2 SkemaFarah Shafiqah AmiruddinNo ratings yet
- DNA, RNA and Protein SynthesisDocument74 pagesDNA, RNA and Protein SynthesisNabilah Rizky Khairunnisa100% (1)
- Biology A (Salters Nuffield) : Pearson Edexcel GCE Level 3 GCEDocument28 pagesBiology A (Salters Nuffield) : Pearson Edexcel GCE Level 3 GCEjNo ratings yet
- Science 10 3rd Quarter Exam - PDF - Messenger Rna - RibosomeDocument1 pageScience 10 3rd Quarter Exam - PDF - Messenger Rna - RibosomeKedir MohammedNo ratings yet
- Organoid Culture HandbookDocument40 pagesOrganoid Culture HandbookGabriella Panuccio100% (1)
- An Algorithmic Immunohistochemical Approach To Define Tumor Type and Assign Site of OriginDocument50 pagesAn Algorithmic Immunohistochemical Approach To Define Tumor Type and Assign Site of OriginCristyan Quintero Múnera100% (1)
- Eals - Urey and MillerDocument7 pagesEals - Urey and MillerKathleen Claire MontefalcoNo ratings yet
- Seaweed Photosyntesis: Rayfan Akbar Fajarinov ViiiDocument5 pagesSeaweed Photosyntesis: Rayfan Akbar Fajarinov ViiiAgus Adi PutrawanNo ratings yet
- Teachers Sec MathematicsDocument21 pagesTeachers Sec MathematicsPRC BaguioNo ratings yet
- Syllabus Breakup - Class 10 (Biology)Document2 pagesSyllabus Breakup - Class 10 (Biology)AaraNo ratings yet
- 4th Q Science 8 DLLDocument6 pages4th Q Science 8 DLLJanecil A. BonzaNo ratings yet
- Searchin Stripped CloverDocument7 pagesSearchin Stripped Clovernacho urbina rodriguezNo ratings yet
- Spore Walkthrough GuideDocument210 pagesSpore Walkthrough GuideJayson BemisNo ratings yet
- GAHUM, JAMAICA P. STS - Module8 - Answer SheetDocument5 pagesGAHUM, JAMAICA P. STS - Module8 - Answer SheetJamaica GahumNo ratings yet
- Life Is CellularDocument15 pagesLife Is Cellularapi-240096234No ratings yet
- Trial SPM SBP 2010 Biology Marking SchemeDocument24 pagesTrial SPM SBP 2010 Biology Marking SchemeMr.D-SIM100% (6)
- Human Biology: Suzanne Wakim & Mandeep GrewalDocument731 pagesHuman Biology: Suzanne Wakim & Mandeep GrewalevertNo ratings yet
- Metabolic Comunication ExerciseDocument12 pagesMetabolic Comunication ExerciseKevin VillaNo ratings yet
- CV - Dr. AlokDocument16 pagesCV - Dr. Alokalok1503No ratings yet
- Dav Public School, Pokhariput, Bhubaneswar Practice Test CL As S - 12 Sub: BiologyDocument6 pagesDav Public School, Pokhariput, Bhubaneswar Practice Test CL As S - 12 Sub: BiologyTushar SinghNo ratings yet
- Creatural Fictions - Human-Animal Relationships in Twentieth - and Twenty-First-Century Literature (David Herman) PDFDocument285 pagesCreatural Fictions - Human-Animal Relationships in Twentieth - and Twenty-First-Century Literature (David Herman) PDFÁdám T. Bogár100% (1)
- Cauliflower Mosaic VirusDocument15 pagesCauliflower Mosaic Virussivagurunathan.P100% (5)
- CK-12 Biology Chapter 25 WorksheetsDocument32 pagesCK-12 Biology Chapter 25 WorksheetsDEEBANNo ratings yet