You might also like
- Roussel Kozon M2eftDocument9 pagesRoussel Kozon M2eftdaniel naoeNo ratings yet
- Body Size and Host Plant SpecializationDocument14 pagesBody Size and Host Plant Specializationmp2233No ratings yet
- Alberdi Et Al 2020Document8 pagesAlberdi Et Al 2020FernandoNo ratings yet
- Eco and Evo 2011Document15 pagesEco and Evo 2011MarcoNo ratings yet
- Group 1 - Scientific PaperDocument31 pagesGroup 1 - Scientific PaperAnthon OdiNo ratings yet
- American Arachnological SocietyDocument8 pagesAmerican Arachnological SocietyEvan SadnessNo ratings yet
- Layman Et Al. - 2007 - Can Stable Isotope Ratios Provide For Community - Wide Measures of Trophic StructureDocument7 pagesLayman Et Al. - 2007 - Can Stable Isotope Ratios Provide For Community - Wide Measures of Trophic StructureLeandrorvNo ratings yet
- 1 s2.0 S0031018221004211 MainDocument16 pages1 s2.0 S0031018221004211 MainAtiqah AuliaNo ratings yet
- Ee35 0684Document10 pagesEe35 0684fungsional fungsionalNo ratings yet
- HamiltonEtAl 2012Document9 pagesHamiltonEtAl 2012Victor Isaac Perez soteloNo ratings yet
- 10 Mamm A 117%2E1Document9 pages10 Mamm A 117%2E1Romie LoveNo ratings yet
- Species Richness and Spatial Diversity of Spiders Among Fodder CropsDocument11 pagesSpecies Richness and Spatial Diversity of Spiders Among Fodder CropsMd Ashikur RahmanNo ratings yet
- Molecular Ecology - 2013 - Delsuc - Convergence of Gut Microbiomes in Myrmecophagous MammalsDocument17 pagesMolecular Ecology - 2013 - Delsuc - Convergence of Gut Microbiomes in Myrmecophagous MammalsleandrasapucaiasNo ratings yet
- Department of Biology, Princeton University, Princeton, New JerseyDocument3 pagesDepartment of Biology, Princeton University, Princeton, New JerseyIsworo RukmiNo ratings yet
- Aguirre Et Al 2003aDocument12 pagesAguirre Et Al 2003aJ E Fernando CernaNo ratings yet
- Ece3 5109Document9 pagesEce3 5109Isaí ValleNo ratings yet
- Melo Etal 2017 Testing Habitat Amount Hypothesis Biol Conserv PDFDocument12 pagesMelo Etal 2017 Testing Habitat Amount Hypothesis Biol Conserv PDFgbitencourt.bioNo ratings yet
- HAYATI Journal of Biosciences: Ida Ayu Ari Janiawati, Mirza Dikari Kusrini, Ani MardiastutiDocument7 pagesHAYATI Journal of Biosciences: Ida Ayu Ari Janiawati, Mirza Dikari Kusrini, Ani MardiastutiOman SetiyantoNo ratings yet
- Libro Parte 2Document78 pagesLibro Parte 2Misael ZagaNo ratings yet
- Morphological Characteristics of Ant Assemblages (Hymenoptera: Formicidae) Differ Among Contrasting BiomesDocument9 pagesMorphological Characteristics of Ant Assemblages (Hymenoptera: Formicidae) Differ Among Contrasting BiomesSalah brahimNo ratings yet
- Research ProposalDocument14 pagesResearch ProposalHaris MusakhelNo ratings yet
- Evaluating Multiple Arthropod Taxa As Indicators of InvertebrateDocument15 pagesEvaluating Multiple Arthropod Taxa As Indicators of InvertebrateCelular DanielaNo ratings yet
- 1 s2.0 S1470160X21003836 MainDocument11 pages1 s2.0 S1470160X21003836 MainMarcelo Aparecido MarquesNo ratings yet
- Oliveira Et Al. 2019 - Lizard Assemblages On Sandy Coastal PlainsDocument12 pagesOliveira Et Al. 2019 - Lizard Assemblages On Sandy Coastal PlainsJuliane RibeiroNo ratings yet
- Pleistocene Mammal EcometricsDocument49 pagesPleistocene Mammal EcometricsjohnhhayesNo ratings yet
- Art. Carrascal Et Al 2012. What Species-Specific Traits Make A Bird A Better Surrogate of Native SpeciesDocument8 pagesArt. Carrascal Et Al 2012. What Species-Specific Traits Make A Bird A Better Surrogate of Native SpeciesmateusfrancaNo ratings yet
- How Mammalian Predation Contributes To T PDFDocument11 pagesHow Mammalian Predation Contributes To T PDFMiguel90No ratings yet
- Evaluation of The Diet of Didelphis Virginiana in An Urban Area Using Stable IsotopesDocument7 pagesEvaluation of The Diet of Didelphis Virginiana in An Urban Area Using Stable Isotopeseyuem28No ratings yet
- Ornelas 2018Document13 pagesOrnelas 2018Nelson AtencioNo ratings yet
- Tropical Forests Are Not Flat: How Mountains Affect Herbivore DiversityDocument10 pagesTropical Forests Are Not Flat: How Mountains Affect Herbivore DiversityMarina Trillo GabaldonNo ratings yet
- Wiley, Ecological Society of America EcologyDocument24 pagesWiley, Ecological Society of America EcologyTatiana LemosNo ratings yet
- Lizard Home Ranges Revisited: Effects of Sex, Body Size, Diet, Habitat, and PhylogenyDocument16 pagesLizard Home Ranges Revisited: Effects of Sex, Body Size, Diet, Habitat, and PhylogenyEric HoNo ratings yet
- Hossie and Murray 2010Document11 pagesHossie and Murray 2010Dennis MurrayNo ratings yet
- May (1994)Document8 pagesMay (1994)star warsNo ratings yet
- Freestone Et Al 2021 Ecology Stronger Predation Intensity Impact On Prey Communities in The TropicsDocument11 pagesFreestone Et Al 2021 Ecology Stronger Predation Intensity Impact On Prey Communities in The TropicsMary Jane AtienzaNo ratings yet
- Bainard Et Al 2020. Genome Size and Endopolyploidy Evolution Across The Moss PhylogenyDocument13 pagesBainard Et Al 2020. Genome Size and Endopolyploidy Evolution Across The Moss PhylogenylaurachalupNo ratings yet
- Ferguson Joly 2002Document9 pagesFerguson Joly 2002Wendy BautistaNo ratings yet
- Journal of Applied Ecology - 2023 - Kemp - Changes in Trophic Ecology of Mobile Predators in Response To RainforestDocument10 pagesJournal of Applied Ecology - 2023 - Kemp - Changes in Trophic Ecology of Mobile Predators in Response To RainforestMuneeba MukhtarNo ratings yet
- Diversity of The Families of Coleoptera Captured With Pitfall Traps in Five Different Environments in Santa Maria, RS, BrazilDocument10 pagesDiversity of The Families of Coleoptera Captured With Pitfall Traps in Five Different Environments in Santa Maria, RS, BrazilSarai Olmedo CruzNo ratings yet
- Agriculture, Ecosystems and Environment: A B C A A D 1 A 1Document8 pagesAgriculture, Ecosystems and Environment: A B C A A D 1 A 1Evan SadnessNo ratings yet
- Menge & Menge, 1974Document22 pagesMenge & Menge, 1974Nataly SlivakNo ratings yet
- Macroecology of Animal Parasitism: Maxim V. VinarskiDocument26 pagesMacroecology of Animal Parasitism: Maxim V. VinarskiNeheLhieNo ratings yet
- Community Structure and Abundance of Small Rodents at The Wave Front of Agroforestry and Forest in Alto Beni, Bolivia - 2020Document10 pagesCommunity Structure and Abundance of Small Rodents at The Wave Front of Agroforestry and Forest in Alto Beni, Bolivia - 2020Abel Tome CaetanoNo ratings yet
- Functional Diversity of Crustacean Zooplankton Communities Towards A Trait-Based Classification 2007 BARNETT Marcado PdfviewDocument18 pagesFunctional Diversity of Crustacean Zooplankton Communities Towards A Trait-Based Classification 2007 BARNETT Marcado PdfviewrgelabertNo ratings yet
- Colston Et Al-2016-Molecular Ecology2222Document25 pagesColston Et Al-2016-Molecular Ecology2222Diana CalderaNo ratings yet
- Comparative Trophic Ecology of Two Sympatric Canids in The Brazilian PampaDocument8 pagesComparative Trophic Ecology of Two Sympatric Canids in The Brazilian PampaLucìa SolerNo ratings yet
- Fernandes Et Al., 2016 - Role of Macrophyte Life Forms in Driving Periphytic...Document8 pagesFernandes Et Al., 2016 - Role of Macrophyte Life Forms in Driving Periphytic...Ubirajara FernandesNo ratings yet
- 418 Article Essay 1Document6 pages418 Article Essay 1Emma ArmitageNo ratings yet
- A Comparative Diversity Analysis of Soil Nematodes of Rajouri, Jammu and Kashmir, IndiaDocument14 pagesA Comparative Diversity Analysis of Soil Nematodes of Rajouri, Jammu and Kashmir, IndiaDr Sharique AliNo ratings yet
- Teoria Nicho ArticuloDocument7 pagesTeoria Nicho ArticuloANA LUCIA CORTES LOAIZANo ratings yet
- GJESM Volume 7 Issue 3 Pages 473-484Document12 pagesGJESM Volume 7 Issue 3 Pages 473-484GJESMNo ratings yet
- Ferruginous Hawk Movements Respond Predictably To Intra Annual Variation But Unexpectedly To Anthropogenic Habitats - Original Article (Habitat Use)Document13 pagesFerruginous Hawk Movements Respond Predictably To Intra Annual Variation But Unexpectedly To Anthropogenic Habitats - Original Article (Habitat Use)buccaneerfosterNo ratings yet
- Frank2017 PDFDocument13 pagesFrank2017 PDFNicolás NovoaNo ratings yet
- British Ecological Society Journal of Animal EcologyDocument10 pagesBritish Ecological Society Journal of Animal EcologyDennis Nabor Muñoz, RN,RMNo ratings yet
- Symbiotic InteractionsDocument18 pagesSymbiotic InteractionsJotha-OriginNo ratings yet
- Sales Et Al 2011 - Feeding Ecology of Ameiva AmeivaDocument10 pagesSales Et Al 2011 - Feeding Ecology of Ameiva AmeivaRaul SalesNo ratings yet
- Correlations Between Lizard Cranial Shape and Diet: A Quantitative, Phylogenetically Informed AnalysisDocument34 pagesCorrelations Between Lizard Cranial Shape and Diet: A Quantitative, Phylogenetically Informed Analysisalexandra gonzalez castilloNo ratings yet
- Functional Traits of Seeds Dispersed Through Endozoochory by Native Forest UngulatesDocument12 pagesFunctional Traits of Seeds Dispersed Through Endozoochory by Native Forest UngulatesAMANDA RAMOSNo ratings yet
- Variation of Butterfly Diet Breadth in Relation To Host-Plant Predictability: Results From Two FaunasDocument17 pagesVariation of Butterfly Diet Breadth in Relation To Host-Plant Predictability: Results From Two FaunasKellyta RodriguezNo ratings yet
- Effects of Resource Distribution on Animal Plant InteractionsFrom EverandEffects of Resource Distribution on Animal Plant InteractionsNo ratings yet
- New Multidimensional Functional Diversity Indices For A Multifaceted Framework in Functional EcologyDocument12 pagesNew Multidimensional Functional Diversity Indices For A Multifaceted Framework in Functional EcologyDaniel Florez TellezNo ratings yet
- Biotropica - 2015 - Schuldt - Tree Species Richness Strengthens Relationships Between Ants and The Functional CompositionDocument8 pagesBiotropica - 2015 - Schuldt - Tree Species Richness Strengthens Relationships Between Ants and The Functional CompositionDaniel Ramos GutierrezNo ratings yet
- Ecography - 2005 - Tsai - Habitat Management by Aboriginals Promotes High Spider Diversity On An Asian Tropical IslandDocument11 pagesEcography - 2005 - Tsai - Habitat Management by Aboriginals Promotes High Spider Diversity On An Asian Tropical IslandDaniel Ramos GutierrezNo ratings yet
- Austral Ecology - 2017 - Barton - Environmental and Spatial Drivers of Spider Diversity at Contrasting MicrohabitatsDocument11 pagesAustral Ecology - 2017 - Barton - Environmental and Spatial Drivers of Spider Diversity at Contrasting MicrohabitatsDaniel Ramos GutierrezNo ratings yet
- CP13-75 1Document7 pagesCP13-75 1Daniel Ramos GutierrezNo ratings yet
- Phylogenetics MygalomorphidaeDocument18 pagesPhylogenetics MygalomorphidaeDaniel Ramos GutierrezNo ratings yet
- Divergence Estimate Chelicerata Mitochon PhylogenyDocument18 pagesDivergence Estimate Chelicerata Mitochon PhylogenyDaniel Ramos GutierrezNo ratings yet
- 2010 Spider Genitalia Chapter Leonard-Cordoba Ch12 DDocument36 pages2010 Spider Genitalia Chapter Leonard-Cordoba Ch12 DDaniel Ramos GutierrezNo ratings yet
- On Some New Genera and Species of Araneida: O P-CambridgeDocument21 pagesOn Some New Genera and Species of Araneida: O P-CambridgeDaniel Ramos GutierrezNo ratings yet
- 7th Sem ClassDocument23 pages7th Sem ClassSatish kumarNo ratings yet
- Rainforest Deforestation Sudent WorksheetDocument2 pagesRainforest Deforestation Sudent WorksheetAlejandro Posada PenagosNo ratings yet
- Speech TextDocument13 pagesSpeech Textdeepti_singh_ntlNo ratings yet
- Biodiversity SangliDocument7 pagesBiodiversity SangliAshish DeotaleNo ratings yet
- Strong Runs Winter 2020Document24 pagesStrong Runs Winter 2020MarkNo ratings yet
- TWCS-I Project-Proposal-2021-2022 - (SPG)Document6 pagesTWCS-I Project-Proposal-2021-2022 - (SPG)Elsie ReyesNo ratings yet
- Unit Vi - Towards A Sustainable WorldDocument9 pagesUnit Vi - Towards A Sustainable WorldAngela Daphnie ToribioNo ratings yet
- Swiss Ecological Scarcity Method: The New Version 2006: January 2006Document5 pagesSwiss Ecological Scarcity Method: The New Version 2006: January 2006Delmy GonzálezNo ratings yet
- NCERT - Tricks Organisms and Populations (501551)Document4 pagesNCERT - Tricks Organisms and Populations (501551)Jeevan narutoNo ratings yet
- BT Anh 12 Thí Điểm Unit 6 - KEYDocument16 pagesBT Anh 12 Thí Điểm Unit 6 - KEYHera TrinhNo ratings yet
- Reaction Paper About Care For Our EnvironmentDocument8 pagesReaction Paper About Care For Our EnvironmentJamesNo ratings yet
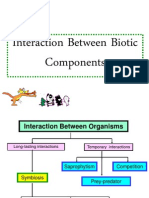
- 01 Interaction Between Biotic ComponentsDocument21 pages01 Interaction Between Biotic ComponentsSharmini Rajagopal100% (1)
- Water$ave Hydro-Tac CatalogueDocument2 pagesWater$ave Hydro-Tac CataloguePolymer InnovationsNo ratings yet
- Pulikulam Cattle Approved As Indigenous BreedDocument2 pagesPulikulam Cattle Approved As Indigenous BreedS.N.RajasekaranNo ratings yet
- Kanha National ParkDocument7 pagesKanha National Parkdeepam0187100% (1)
- General Education Pointers To Review: 1. ENGLISH - 25% (Of The Whole General Education Part of The Exam)Document6 pagesGeneral Education Pointers To Review: 1. ENGLISH - 25% (Of The Whole General Education Part of The Exam)Rocky YapNo ratings yet
- Tropical ClimatesDocument15 pagesTropical ClimatesKeshavSeeamNo ratings yet
- Blanqueamineto Coral RapanuiDocument18 pagesBlanqueamineto Coral RapanuiMare Del PacifícoNo ratings yet
- 7334-Article Text-25839-1-10-20200529Document16 pages7334-Article Text-25839-1-10-20200529ilhamNo ratings yet
- OAI551-Environment and Agriculture Question BankDocument9 pagesOAI551-Environment and Agriculture Question BankMohamed Raees100% (1)
- Unit-2-Natural ResourcesDocument10 pagesUnit-2-Natural ResourcesJenx TechzNo ratings yet
- Soil Erosion EssayDocument6 pagesSoil Erosion Essaylwfdwwwhd100% (2)
- Fall 2003 Muir Heritage Land Trust NewsletterDocument12 pagesFall 2003 Muir Heritage Land Trust NewsletterMuir Heritage Land TrustNo ratings yet
- Eco Tourism DR Meeta Nihalani Kajol DhootDocument19 pagesEco Tourism DR Meeta Nihalani Kajol Dhootdr meetaNo ratings yet
- BMB-UWM 2017 - PaPIS &NPDinput - Sept 23 (2) - May 4, 2017Document44 pagesBMB-UWM 2017 - PaPIS &NPDinput - Sept 23 (2) - May 4, 2017cenro staritaNo ratings yet
- Bba Evs Question BNKDocument2 pagesBba Evs Question BNKHarnoor Singh -No ratings yet
- Various Elements That May Be Exposed To HazardsDocument35 pagesVarious Elements That May Be Exposed To Hazardsjoel50% (2)
- Lab ReportDocument6 pagesLab Reportapi-375366961No ratings yet
- Bird-Friendly Trees EGDocument13 pagesBird-Friendly Trees EGSylvia T RamosNo ratings yet