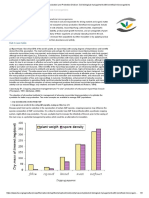
You might also like
- Environmental Impact Assessment: Final Report After Public HearingDocument554 pagesEnvironmental Impact Assessment: Final Report After Public HearingRishabh100% (3)
- VERMICOMPOSTINGDocument41 pagesVERMICOMPOSTINGNeha Srivastava100% (1)
- Azolla Preparation PDFDocument2 pagesAzolla Preparation PDFLloydie Lopez100% (2)
- Culture of Rotifer With CommentsDocument20 pagesCulture of Rotifer With CommentsJerick Christian P Dagdagan0% (1)
- Plant Growth Promoting Rhizobacteria in Crop Protection and ChallengesDocument29 pagesPlant Growth Promoting Rhizobacteria in Crop Protection and ChallengesTrái Tim BăngNo ratings yet
- Beneficial Bacteria of Agricultural ImportanceDocument12 pagesBeneficial Bacteria of Agricultural ImportanceXevin MoreiraNo ratings yet
- An and HiDocument5 pagesAn and HijjprianNo ratings yet
- Formulations of Bacillus Spp. and Pseudomonas Root Rot Caused by Fusarium SolaniDocument6 pagesFormulations of Bacillus Spp. and Pseudomonas Root Rot Caused by Fusarium SolaniNelver MorenoNo ratings yet
- Impact of Antifungals Producing Rhizobacteria On The Performance of Vigna Radiata in The Presence of Arbuscular Mycorrhizal FungiDocument12 pagesImpact of Antifungals Producing Rhizobacteria On The Performance of Vigna Radiata in The Presence of Arbuscular Mycorrhizal FungimetbijayNo ratings yet
- Cisgenesis and Intragenesis: New Tools For Improving CropsDocument10 pagesCisgenesis and Intragenesis: New Tools For Improving CropsgeraldineChrisNo ratings yet
- Contreras Cornejo 2017Document9 pagesContreras Cornejo 2017Karen Avalos VelaNo ratings yet
- Bacteria For Plant Growth Promotion andDocument34 pagesBacteria For Plant Growth Promotion andrahimmarriNo ratings yet
- Biofumigation: A Potential Aspect For Suppression of Plant-Parasitic NematodesDocument7 pagesBiofumigation: A Potential Aspect For Suppression of Plant-Parasitic NematodesIJEAB JournalNo ratings yet
- Plant Production and Protection Division - Soil Biological Management With Beneficial MicroorganismsDocument3 pagesPlant Production and Protection Division - Soil Biological Management With Beneficial MicroorganismsCyberSquare IndiaNo ratings yet
- Cucumber-Bhaskar ChatterjeeDocument15 pagesCucumber-Bhaskar ChatterjeeBhaskar ChatterjeeNo ratings yet
- Trangenic TrigoDocument11 pagesTrangenic TrigoCesarRamNo ratings yet
- Soil Beneficial Bacteria and Their Role in Plant GrowthDocument20 pagesSoil Beneficial Bacteria and Their Role in Plant GrowthKhairilanuar Mohd HanimNo ratings yet
- Application of Fluorescent Pseudomonads Inoculant Formulations On Vigna Mungo Through Field TrialDocument5 pagesApplication of Fluorescent Pseudomonads Inoculant Formulations On Vigna Mungo Through Field TrialcarloscarinelliNo ratings yet
- Salt StressDocument5 pagesSalt StressMahtab KhurshidNo ratings yet
- Gdanetz and Trail PBIOMES-05!17!0023-RDocument11 pagesGdanetz and Trail PBIOMES-05!17!0023-RCandysoup101No ratings yet
- Seed Biopriming PGPR IntrodDocument14 pagesSeed Biopriming PGPR IntrodMohammad Mosharraf HossainNo ratings yet
- Evaluation of Rhizosphere Bacterial Antagonists For Their Potential To BioprotectDocument7 pagesEvaluation of Rhizosphere Bacterial Antagonists For Their Potential To BioprotectHvzem MohammedNo ratings yet
- Reviewed ArticleDocument6 pagesReviewed ArticleWaleed RamzanNo ratings yet
- LupinDocument16 pagesLupinYassertahlawyNo ratings yet
- Chapter Genetically Modified CropsDocument10 pagesChapter Genetically Modified CropsMazhar FarNo ratings yet
- ACC Deaminase MethodsDocument6 pagesACC Deaminase MethodsMala MajumdarNo ratings yet
- PT 3163 66o With Cover Page v2Document5 pagesPT 3163 66o With Cover Page v2Andreina ZamoraNo ratings yet
- Art:10.1007/s11101 006 9032 2Document35 pagesArt:10.1007/s11101 006 9032 2Caroline LopesNo ratings yet
- 2 Ijasrjun20182Document26 pages2 Ijasrjun20182TJPRC PublicationsNo ratings yet
- 10 1007@s005720050147Document8 pages10 1007@s005720050147arthurgladysntabalaNo ratings yet
- 2015 Article 327Document11 pages2015 Article 327Dicson Sánchez AbadNo ratings yet
- Torres 2003Document6 pagesTorres 2003Thu UyênNo ratings yet
- Toppo and Nail 2015Document9 pagesToppo and Nail 2015Alan Rivera IbarraNo ratings yet
- tmpD493 TMPDocument11 pagestmpD493 TMPFrontiersNo ratings yet
- Simsek-Ersahin f1Document7 pagesSimsek-Ersahin f1YurdagulNo ratings yet
- Management of Bacterial Wilt (Ralstonia Solanacearum) of Potato: Focus On Natural Bioactive CompoundsDocument20 pagesManagement of Bacterial Wilt (Ralstonia Solanacearum) of Potato: Focus On Natural Bioactive Compoundsdawit gNo ratings yet
- tmp43EC TMPDocument7 pagestmp43EC TMPFrontiersNo ratings yet
- tmpAEC8 TMPDocument12 pagestmpAEC8 TMPFrontiersNo ratings yet
- Effects of The Co Inoculation With Saprobe and Mycorrhizal Fungi On Vaccinium Corymbosum Growth and Some Soil Enzymatic ActivitiesDocument12 pagesEffects of The Co Inoculation With Saprobe and Mycorrhizal Fungi On Vaccinium Corymbosum Growth and Some Soil Enzymatic Activitiesjadofo6No ratings yet
- Agri Sci - Ijasr-Mass Production of Biofertilizer - Pseudomonas FluorescensDocument4 pagesAgri Sci - Ijasr-Mass Production of Biofertilizer - Pseudomonas FluorescensTJPRC PublicationsNo ratings yet
- Bakhshi (2020)Document7 pagesBakhshi (2020)Gleice LimaNo ratings yet
- tmpEDC0 TMPDocument11 pagestmpEDC0 TMPFrontiersNo ratings yet
- 09 Chapter 2Document51 pages09 Chapter 2Anonymous OP6R1ZSNo ratings yet
- Physiological, Nutritional, and Biochemical Bases of Corn Resistance To Foliage-Feeding Fall ArmywormDocument10 pagesPhysiological, Nutritional, and Biochemical Bases of Corn Resistance To Foliage-Feeding Fall ArmywormSilvio FaveroNo ratings yet
- Plant Growth Promoting Rhizobacteria and Sustainable Agriculture: A ReviewDocument6 pagesPlant Growth Promoting Rhizobacteria and Sustainable Agriculture: A ReviewMohammad Mosharraf HossainNo ratings yet
- Utilization of Arbuscular Micorrhizal Fungi To Control Fusarium Wilt of TomatoesDocument6 pagesUtilization of Arbuscular Micorrhizal Fungi To Control Fusarium Wilt of TomatoeswafiyyatunnufusNo ratings yet
- Biological Control of NematodesDocument17 pagesBiological Control of NematodesYara Othman Abdel RaoufNo ratings yet
- Fpls 13 897668Document14 pagesFpls 13 897668clingoNo ratings yet
- Microorganisms 08 01329 v2Document18 pagesMicroorganisms 08 01329 v2Muhamad AmirulNo ratings yet
- Organic Production of Medicinal, Aromatic and Dye-Yielding Plants (Madps) - With Inputs From FRLHTDocument7 pagesOrganic Production of Medicinal, Aromatic and Dye-Yielding Plants (Madps) - With Inputs From FRLHTanon-189290No ratings yet
- Challenges and Opportunities in Implementing Allelopathy For Natural Weed ManagementDocument32 pagesChallenges and Opportunities in Implementing Allelopathy For Natural Weed Managementbrkica2011No ratings yet
- Jayraj Synposis Okra 1Document24 pagesJayraj Synposis Okra 1MOHIT SomaniNo ratings yet
- Biological Control of Rice Bacterial Blight by Plant Associated BacteriaDocument10 pagesBiological Control of Rice Bacterial Blight by Plant Associated BacteriaSufian TapadarNo ratings yet
- Preliminary Investigations On Selection of Synergistic Halotolerant Plant Growth Promoting Rhizobacteria For Inducing Salinity Tolerance in WheatDocument11 pagesPreliminary Investigations On Selection of Synergistic Halotolerant Plant Growth Promoting Rhizobacteria For Inducing Salinity Tolerance in WheatTahiri AbdelilahNo ratings yet
- Kumar Et Al 2013 Evaluating The Plant Growth Promoting AbilityDocument8 pagesKumar Et Al 2013 Evaluating The Plant Growth Promoting AbilitykumarmicroNo ratings yet
- Biocontrol PDFDocument6 pagesBiocontrol PDFWesh De GeminosNo ratings yet
- Chapter 2 - Q3 Group 6Document2 pagesChapter 2 - Q3 Group 6Gian Carlo M. AquinoNo ratings yet
- Effects of Azospirillum Bacteria and Gibberellin HormoneDocument8 pagesEffects of Azospirillum Bacteria and Gibberellin HormoneOliver TalipNo ratings yet
- Gupta Et Al 2012Document12 pagesGupta Et Al 2012rg1326No ratings yet
- 1 s2.0 S094450131300116X MainDocument7 pages1 s2.0 S094450131300116X MainJorge CorderoNo ratings yet
- 73-82 (4627)Document10 pages73-82 (4627)Afif FebrianNo ratings yet
- Plant Growth Promoting RhizobiaDocument23 pagesPlant Growth Promoting RhizobiadanyjorgeNo ratings yet
- Lant Rotection: How To Diagnose Plant ProblemsDocument224 pagesLant Rotection: How To Diagnose Plant Problemsكركشندي أفندي Karkshndy afndyNo ratings yet
- 2021 01 20 427489v1 FullDocument26 pages2021 01 20 427489v1 Fullكركشندي أفندي Karkshndy afndyNo ratings yet
- ADR PEG and Starch Based ThickenersDocument1 pageADR PEG and Starch Based Thickenersكركشندي أفندي Karkshndy afndyNo ratings yet
- ملخص 200 كلمة انجليزيDocument1 pageملخص 200 كلمة انجليزيكركشندي أفندي Karkshndy afndyNo ratings yet
- A Summary of The Sugar RevolutionDocument2 pagesA Summary of The Sugar RevolutionConrad SawyersNo ratings yet
- Mineral Nutrition and Suppression of Plant Disease - Elmer2014Document14 pagesMineral Nutrition and Suppression of Plant Disease - Elmer2014Dario BaronaNo ratings yet
- Indigenous Filipinos During American Colonization PeriodDocument5 pagesIndigenous Filipinos During American Colonization PeriodHowell AragonNo ratings yet
- Serengeti Shall Not Die PDFDocument20 pagesSerengeti Shall Not Die PDFTeklehaymanot GeremeskelNo ratings yet
- Potato ProjectDocument10 pagesPotato Projectkillerfox70No ratings yet
- 117th Issue New DawnDocument16 pages117th Issue New DawnUmmah_KenyaNo ratings yet
- A House Not Made With HandsDocument125 pagesA House Not Made With HandsRosy ColeNo ratings yet
- TestDocument27 pagesTestHoàng ThọNo ratings yet
- 4 Plant Disease General ConceptsDocument35 pages4 Plant Disease General ConceptsTathagata Ray ChaudhuriNo ratings yet
- Dairy Development 4Document39 pagesDairy Development 4Ancy RajNo ratings yet
- Bayer A HistoryDocument6 pagesBayer A HistorygokundiNo ratings yet
- 2001 A Household Analysis of Huastec MayDocument13 pages2001 A Household Analysis of Huastec MayChad MinottNo ratings yet
- CARPER RA 9700 - Comprehensive Agrarian Reform Program Extension With ReformsDocument13 pagesCARPER RA 9700 - Comprehensive Agrarian Reform Program Extension With ReformsVon Darryl Gerale Mabuna100% (2)
- Dizzy With SuccessDocument5 pagesDizzy With Successapi-3851760No ratings yet
- Chocolate SlaveryDocument5 pagesChocolate SlaveryAnnaHernandezNo ratings yet
- 4house of Flameballah Wedo: Apells and Ebbis Part 4 PDFDocument3 pages4house of Flameballah Wedo: Apells and Ebbis Part 4 PDFFelipe ArandaNo ratings yet
- SocialScience9th FNL PDFDocument206 pagesSocialScience9th FNL PDFAryanNo ratings yet
- Profile: 270 Lbs. Man Who Has Always Had A Hard Time Gaining WeightDocument10 pagesProfile: 270 Lbs. Man Who Has Always Had A Hard Time Gaining WeightYuuki XDNo ratings yet
- Asociación de Productores Cacaoteros y Cafetaleros de Amazonas AprocamDocument3 pagesAsociación de Productores Cacaoteros y Cafetaleros de Amazonas AprocamEric JaraNo ratings yet
- Feasibility StudiesDocument3 pagesFeasibility StudiesKelvin John Sajulan Cabaniero50% (2)
- FULL DOCUMENT - Organic FoodDocument9 pagesFULL DOCUMENT - Organic FoodNeerajNo ratings yet
- Narrative Report Part 2Document30 pagesNarrative Report Part 2Randy FernandezNo ratings yet
- 07 Chapter 2Document146 pages07 Chapter 2tpgggkNo ratings yet
- GarinMap 20160721 PDFDocument2 pagesGarinMap 20160721 PDFjoker8832No ratings yet
- Roman Candle Middleton CandleDocument4 pagesRoman Candle Middleton Candlesupport_local_flavorNo ratings yet
- Peak Oil - WDocument26 pagesPeak Oil - Wvincent brazilNo ratings yet