You might also like
- Design On Slab On GradeDocument4 pagesDesign On Slab On GradeAhmad Mensa100% (1)
- 16.1 HVAC Master Cheat Sheet Document Revised PDFDocument25 pages16.1 HVAC Master Cheat Sheet Document Revised PDFDiana BarchukNo ratings yet
- Oil Tanker PipingDocument38 pagesOil Tanker PipingnikhilNo ratings yet
- (2022) Müller Et Al. (Stenoscelida Aurantiacus)Document20 pages(2022) Müller Et Al. (Stenoscelida Aurantiacus)mauricio.garciaNo ratings yet
- Vermicomposting in The PhilippinesDocument23 pagesVermicomposting in The PhilippinesElisha TajanNo ratings yet
- Allen Press and American Society of Ichthyologists and Herpetologists (ASIH) Are Collaborating With JSTOR To Digitize, CopeiaDocument9 pagesAllen Press and American Society of Ichthyologists and Herpetologists (ASIH) Are Collaborating With JSTOR To Digitize, CopeiaEsther GnrNo ratings yet
- Three New Species of Remaneicaris Jakobi 1972 CopeDocument29 pagesThree New Species of Remaneicaris Jakobi 1972 CopeEdinaldo Nelson Dos Santos SilvaNo ratings yet
- DownloadDocument9 pagesDownloadEduarda SerraNo ratings yet
- Characterization and Taxonomic Key For Tadpoles From The Northwestern Region of São Paulo State, BrazilDocument27 pagesCharacterization and Taxonomic Key For Tadpoles From The Northwestern Region of São Paulo State, Braziljonho2006No ratings yet
- Deforeletal 2021melanophryniscusDocument27 pagesDeforeletal 2021melanophryniscusjose.1711717678No ratings yet
- Artículo de RenacuajosDocument9 pagesArtículo de RenacuajosAndres Felipe BermudezNo ratings yet
- Notes On The Taxonomy of Mountain Viscachas of The Genus: Lagidium Meyen 1833 (Rodentia: Chinchillidae)Document8 pagesNotes On The Taxonomy of Mountain Viscachas of The Genus: Lagidium Meyen 1833 (Rodentia: Chinchillidae)Roger Barboza CastroNo ratings yet
- Martinsetal2016 AncylothrixDocument11 pagesMartinsetal2016 AncylothrixHigor LessaNo ratings yet
- Bradypus Journal of Mammalian Evolution-2019Document24 pagesBradypus Journal of Mammalian Evolution-2019Manuel RuizNo ratings yet
- Pratigi AristeuiDocument17 pagesPratigi AristeuiBeteapoilsNo ratings yet
- Bicentenariella Sinuosa Serranid Fishes New Species 2021 Parecida A Monogeneo de Cichlasoma IstlanumDocument11 pagesBicentenariella Sinuosa Serranid Fishes New Species 2021 Parecida A Monogeneo de Cichlasoma IstlanumBelem FloresNo ratings yet
- 2358 2936 Nau 29 E2021015Document27 pages2358 2936 Nau 29 E2021015pcorgoNo ratings yet
- Natural History of Phyllomedusa Centralis (Anura Hylidae Phyllomedusinae)Document9 pagesNatural History of Phyllomedusa Centralis (Anura Hylidae Phyllomedusinae)Bivek gautamNo ratings yet
- 036 Chiasmocleis in The Atlantic Rain Forest CruzCaramaschiIzecksohn 1997 PDFDocument24 pages036 Chiasmocleis in The Atlantic Rain Forest CruzCaramaschiIzecksohn 1997 PDFAna Karolina MorenoNo ratings yet
- AABC V83n1a20 PepesuchusDocument11 pagesAABC V83n1a20 PepesuchusKanekoNo ratings yet
- New Species of Melanophryniscus (Anura: Bufonidae) From Rio Grande Do Sul, Southern BrazilDocument6 pagesNew Species of Melanophryniscus (Anura: Bufonidae) From Rio Grande Do Sul, Southern BrazilAna AriasNo ratings yet
- CheckList Article 18054 en 1Document3 pagesCheckList Article 18054 en 1Gamer RafaNo ratings yet
- Description of Larva, Pupa, and Genitalia of Hybosa Acutangula Spaeth, 1913Document7 pagesDescription of Larva, Pupa, and Genitalia of Hybosa Acutangula Spaeth, 1913lucasdisarevNo ratings yet
- Phylogenetic Relationships of Calomys Sorellus Complex (Rodentia: Cricetidae), With The Description of Two New SpeciesDocument24 pagesPhylogenetic Relationships of Calomys Sorellus Complex (Rodentia: Cricetidae), With The Description of Two New SpeciesRudy ConchaNo ratings yet
- Condez Etal2014 Bcrispus PDFDocument12 pagesCondez Etal2014 Bcrispus PDFvanessa e bruno becaciciNo ratings yet
- Eterovick and Sazima 1998Document7 pagesEterovick and Sazima 1998abraaoabdaladinizNo ratings yet
- Biogeografia - Arañas en ColombiaDocument49 pagesBiogeografia - Arañas en ColombiaLilia SierraNo ratings yet
- Rando Et Al. 2016Document16 pagesRando Et Al. 2016Isabel PinheiroNo ratings yet
- Dos Nuevas Especies de PhyllodactylusDocument12 pagesDos Nuevas Especies de PhyllodactylusLuis F. GarcíaNo ratings yet
- Taborda Et Al., 2023 Riojasuchus BiomechanicsDocument11 pagesTaborda Et Al., 2023 Riojasuchus Biomechanicsmauricio.garciaNo ratings yet
- Second World Record For Two Calvatia Species (Agaricaceae: Basidiomycota)Document6 pagesSecond World Record For Two Calvatia Species (Agaricaceae: Basidiomycota)Josenildo Ferreira FerreiraNo ratings yet
- Nogueira 2012Document14 pagesNogueira 2012Anonymous VqDPRFNo ratings yet
- Suarezetal 2012 Mast Neot PeropteryxleucopteraypallidopterafromColombiaDocument8 pagesSuarezetal 2012 Mast Neot PeropteryxleucopteraypallidopterafromColombiajose PomaNo ratings yet
- Aspidoras TaxonomiaDocument159 pagesAspidoras TaxonomiaJeferson Luizi PereiraNo ratings yet
- Amorphinopsis (Halichondrida: Demospongiae) From The Atlantic Ocean, With The Description of A New SpeciesDocument7 pagesAmorphinopsis (Halichondrida: Demospongiae) From The Atlantic Ocean, With The Description of A New SpeciesMara Paola DurangoNo ratings yet
- A New Species of The Genus Stenocercus (Iguania Tropiduridae) From The Peruvian Pacific Coast (Ica Region) PDFDocument15 pagesA New Species of The Genus Stenocercus (Iguania Tropiduridae) From The Peruvian Pacific Coast (Ica Region) PDFRenzo Alessandro Pacherres NietoNo ratings yet
- Lynch.1982. Two New Species of Poison Dart Frogs - Edwardsi&ruiziDocument10 pagesLynch.1982. Two New Species of Poison Dart Frogs - Edwardsi&ruiziRaiza Nathaly Castañeda BonillaNo ratings yet
- PeripatusDocument9 pagesPeripatusPaulo de Tárcio Jr.No ratings yet
- Ramirez, Birindelli, Galetti - 2017 - A New Genus of Anostomidae (Ostariophysi Characiformes) Diversity, Phylogeny and Biogeography BaseDocument16 pagesRamirez, Birindelli, Galetti - 2017 - A New Genus of Anostomidae (Ostariophysi Characiformes) Diversity, Phylogeny and Biogeography BaseAmanda CocovickNo ratings yet
- Revision of The Genus Murunducaris Copepoda HarpacDocument22 pagesRevision of The Genus Murunducaris Copepoda HarpacpcorgoNo ratings yet
- Nematopsis Crassostrea RhizophoraeDocument8 pagesNematopsis Crassostrea RhizophoraeRenato NascimentoNo ratings yet
- Brcko Et Al. (2013) Taxonomy and Distribution of The Salamander Genus Bolitoglossa in Brazilian AmazoniaDocument31 pagesBrcko Et Al. (2013) Taxonomy and Distribution of The Salamander Genus Bolitoglossa in Brazilian AmazoniaUécson SuendelNo ratings yet
- Zootaxa: CorrespondenceDocument5 pagesZootaxa: Correspondencecabruca1No ratings yet
- Rovito (2010) Bolitoglossa ZacapensisDocument12 pagesRovito (2010) Bolitoglossa ZacapensisFLOR DE MARIA MORALES ARROYONo ratings yet
- First Record of Stenochrus Portoricensis Chamberlin, 1922 (Arachnida: Schizomida: Hubbardiidae) For The Pernambuco State, BrazilDocument2 pagesFirst Record of Stenochrus Portoricensis Chamberlin, 1922 (Arachnida: Schizomida: Hubbardiidae) For The Pernambuco State, BrazilAndré LiraNo ratings yet
- Agrellos 2012Document28 pagesAgrellos 2012abrahamNo ratings yet
- Oncidiinae (Orchidaceae) On The Great Curve of The Xingu River, Pará State, BrazilDocument16 pagesOncidiinae (Orchidaceae) On The Great Curve of The Xingu River, Pará State, BrazilFagno AvizNo ratings yet
- Taxonomic Key For The Genera of Elmidae ColeopteraDocument9 pagesTaxonomic Key For The Genera of Elmidae ColeopteraFelipe BarbosaNo ratings yet
- Revision of Philodryas Mattogrossensis With The Revalidation of P. Erlandi (Reptilia: Squamata: Dipsadidae)Document15 pagesRevision of Philodryas Mattogrossensis With The Revalidation of P. Erlandi (Reptilia: Squamata: Dipsadidae)Daniela CerqueiraNo ratings yet
- Morato - 2011 - Bothropoides Jararaca Prey - Herp - ReviewDocument2 pagesMorato - 2011 - Bothropoides Jararaca Prey - Herp - ReviewSérgio Augusto Abrahão MoratoNo ratings yet
- Fernandes Et La. 2019. Synopsis of Terrestrial Isopods (CrustaceaDocument36 pagesFernandes Et La. 2019. Synopsis of Terrestrial Isopods (Crustaceapqp128No ratings yet
- Boldrini Et Al., 2017 (Brasil) BaDocument10 pagesBoldrini Et Al., 2017 (Brasil) Bacarmen villalobosNo ratings yet
- Loreto PeruherpsDocument69 pagesLoreto PeruherpsRousse OrtizNo ratings yet
- 2016 Roque Et Al DISDocument2 pages2016 Roque Et Al DISRosana TidonNo ratings yet
- Nunes de Almeida Et Al 2021Document16 pagesNunes de Almeida Et Al 2021carlitos salazarNo ratings yet
- UntitledDocument18 pagesUntitledGabriel AssunçãoNo ratings yet
- Notes On The Courtship Behavior of Aplastodiscus ADocument9 pagesNotes On The Courtship Behavior of Aplastodiscus ABeatriz BritoNo ratings yet
- El Complejo Sporobolus Indicus (Poaceae, Chloridoideae, Zoysieae) en La ArgentinaDocument12 pagesEl Complejo Sporobolus Indicus (Poaceae, Chloridoideae, Zoysieae) en La ArgentinaGuido BarallaNo ratings yet
- SaimiripublicadoPrimates145 161Document19 pagesSaimiripublicadoPrimates145 161Manuel RuizNo ratings yet
- Ostracodes (Crustacea) From Cananéia-Iguape Estuarine/lagoon System and Geographical Distribution of The Mixohaline Assemblages in Southern and Southeastern BrazilDocument8 pagesOstracodes (Crustacea) From Cananéia-Iguape Estuarine/lagoon System and Geographical Distribution of The Mixohaline Assemblages in Southern and Southeastern Braziljoão carlos CoimbraNo ratings yet
- Lobo, Nenda & Slodki 2012Document15 pagesLobo, Nenda & Slodki 2012leandroraulantoniazziNo ratings yet
- A New Subspecies of Saguinus Fuscicollis Primates CallitrichidaeDocument20 pagesA New Subspecies of Saguinus Fuscicollis Primates CallitrichidaeDaysa AthaydesNo ratings yet
- Barrosasuchus Neuquenianus PDFDocument17 pagesBarrosasuchus Neuquenianus PDFFelipe MesVasNo ratings yet
- EE5630 - EE5660 - Exam - Main - FinalDocument5 pagesEE5630 - EE5660 - Exam - Main - FinalMadalin PetrachiNo ratings yet
- Jco Connections Required For Ess Mss ConfigurationDocument6 pagesJco Connections Required For Ess Mss ConfigurationashukennaNo ratings yet
- 07 Subject Verb AgreementDocument14 pages07 Subject Verb AgreementMehak SrivastavaNo ratings yet
- Example of Delivery NoteDocument1 pageExample of Delivery NoteRuDy RaviNo ratings yet
- KZV Newsletter - JanuaryDocument9 pagesKZV Newsletter - JanuaryHasmik MehranianNo ratings yet
- ABSU Post-UTME Past Questions and Answers For Science Students (Sciences, Medicine, Engineering and Agriculture)Document45 pagesABSU Post-UTME Past Questions and Answers For Science Students (Sciences, Medicine, Engineering and Agriculture)Kelvin Rex100% (1)
- B1 UNIT 7 Flipped Classroom Video WorksheetDocument1 pageB1 UNIT 7 Flipped Classroom Video WorksheetHamza AlibegovicNo ratings yet
- Molasses Tank Draft GTP - 12000MT Net-15000 MT GrossDocument14 pagesMolasses Tank Draft GTP - 12000MT Net-15000 MT GrossStressDyn ConsultantsNo ratings yet
- Advanced Hair Transplant 5Document18 pagesAdvanced Hair Transplant 5Naveed KhanNo ratings yet
- Ra-025142 Civil Engineer Davao 11-2021-With PregnantDocument94 pagesRa-025142 Civil Engineer Davao 11-2021-With PregnantVicent John ParedesNo ratings yet
- Hien - Motivational LetterDocument2 pagesHien - Motivational Letterqhuyvu0% (1)
- The Story of The Blind Baba-AbdallaDocument21 pagesThe Story of The Blind Baba-AbdallaJacob DichosaNo ratings yet
- Auroras Whole Realms Autumn CatalogueDocument50 pagesAuroras Whole Realms Autumn CatalogueDiss onanceNo ratings yet
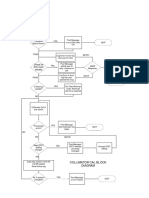
- Check For Dirty Mylar Window: Collimator Cal Block DiagramDocument1 pageCheck For Dirty Mylar Window: Collimator Cal Block DiagramMai Thanh SơnNo ratings yet
- Capital Market LineDocument2 pagesCapital Market LineKaran BhatiaNo ratings yet
- Jessica - Journal (Revisi)Document7 pagesJessica - Journal (Revisi)jessica paulimaNo ratings yet
- Karish Newsletter Issue 4 - DECEMBER 2019 FINALDocument11 pagesKarish Newsletter Issue 4 - DECEMBER 2019 FINALJodan AsiborNo ratings yet
- Untitled DocumentDocument4 pagesUntitled Documentallyssa abrogenaNo ratings yet
- List of Bluesign Approved Chemical Products of TransfarDocument8 pagesList of Bluesign Approved Chemical Products of TransfarMainulHoqueNo ratings yet
- P2 CH 4-8+SM1 Practice Paper - QP3Document12 pagesP2 CH 4-8+SM1 Practice Paper - QP3Thi Hanh Nguyen LeNo ratings yet
- IGCSE Geography - Tourism Case Study BaliDocument9 pagesIGCSE Geography - Tourism Case Study BaliMiguel Oubiña SánchezNo ratings yet
- ARYAN DAA Practical FILE PDFDocument18 pagesARYAN DAA Practical FILE PDFARYAN JAISWALNo ratings yet
- Weiner BibDocument6 pagesWeiner Bibiskandar.hai2226No ratings yet
- Problem Set 6 Fall 2018 With SolutionsDocument5 pagesProblem Set 6 Fall 2018 With SolutionsrickNo ratings yet
- Living With The Himalayan MastersDocument27 pagesLiving With The Himalayan MastersLauren Hughes0% (3)
- 3 Properties of Real Numbers ChartDocument1 page3 Properties of Real Numbers ChartwlochNo ratings yet