You might also like
- Glycogen MetabolismDocument37 pagesGlycogen MetabolismERIAS TENYWANo ratings yet
- 9 GlycogenDocument36 pages9 GlycogenSneha Sagar SharmaNo ratings yet
- Glycogen MetabolismDocument11 pagesGlycogen MetabolismShruti MishraNo ratings yet
- GLYCOGEN METABOLISMDocument63 pagesGLYCOGEN METABOLISMyixecix709No ratings yet
- Glycolysis: The Energy-Extracting PathwayDocument48 pagesGlycolysis: The Energy-Extracting PathwayTeodora MunteanuNo ratings yet
- Glycogen Synthesis 2Document39 pagesGlycogen Synthesis 2CLEMENTNo ratings yet
- Glucolisis & Fermentación Biología VDocument37 pagesGlucolisis & Fermentación Biología Vcamy.acosta.camposNo ratings yet
- Carbohydrates: Answers To QuestionsDocument3 pagesCarbohydrates: Answers To QuestionsGaby de GuzmanNo ratings yet
- Carbohidratos Estructura PDFDocument6 pagesCarbohidratos Estructura PDFPatricia NarvaezNo ratings yet
- Drug elimination (2): Liver metabolismDocument26 pagesDrug elimination (2): Liver metabolismSylvia AngelinaNo ratings yet
- Pentose Phosphate Pathway: Molecular Biochemistry IIDocument27 pagesPentose Phosphate Pathway: Molecular Biochemistry IIElena PavlicNo ratings yet
- Glycolysis: Biochemistry of MetabolismDocument47 pagesGlycolysis: Biochemistry of MetabolismDozdiNo ratings yet
- Carbs: Sources of Energy, Structures & FunctionsDocument28 pagesCarbs: Sources of Energy, Structures & FunctionsKiruthikasweetyNo ratings yet
- Wa0013.Document30 pagesWa0013.VekeMalaNo ratings yet
- GlycolysisDocument44 pagesGlycolysisGilbert TanNo ratings yet
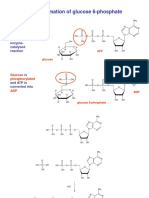
- Formation of Glucose 6-PhosphateDocument2 pagesFormation of Glucose 6-PhosphateSanjeefKumrIINo ratings yet
- 14glucose6 PpsDocument2 pages14glucose6 PpsSanjeefKumrIINo ratings yet
- GlycolosisDocument49 pagesGlycolosismishraravikumar428No ratings yet
- BMB1203 Biochem IIlecture 1Document120 pagesBMB1203 Biochem IIlecture 1Brian Adoka OmdangNo ratings yet
- 3 Activity Carbohyrdates IDocument10 pages3 Activity Carbohyrdates INur SetsuNo ratings yet
- 2022 Edited Chemistry of Carbohydrate 2Document33 pages2022 Edited Chemistry of Carbohydrate 2kel GetanehNo ratings yet
- Available compound as a starting material for organic synthesisDocument10 pagesAvailable compound as a starting material for organic synthesisAfifaNo ratings yet
- Lipid BiosynthesisDocument60 pagesLipid BiosynthesisNdy AgustaviaNo ratings yet
- UNIT-14 BIOMOLECULES 2023Document22 pagesUNIT-14 BIOMOLECULES 2023jagannathanNo ratings yet
- Structure and Function of CarbohydratesDocument27 pagesStructure and Function of CarbohydratesAli SeenaNo ratings yet
- Legaturi glicozidice si structuri glucidice complexeDocument7 pagesLegaturi glicozidice si structuri glucidice complexedeedeeutza14No ratings yet
- Chapter 2 - Carbohydrate MetabolismDocument72 pagesChapter 2 - Carbohydrate MetabolismPJ1-0619 Florence Binti IstemNo ratings yet
- Carbohydrates I: Common Reactions 3: Laboratory ManualDocument10 pagesCarbohydrates I: Common Reactions 3: Laboratory ManualJhet Ardian CoritanaNo ratings yet
- Biochem2 Carbs and LipidsDocument69 pagesBiochem2 Carbs and Lipidstml_19672682No ratings yet
- Phương Trình Con Đư NG EMPDocument2 pagesPhương Trình Con Đư NG EMPDương TháiNo ratings yet
- Experiment 5 Properties of Carbohydrates: Solubility, Reactivity, and Specific RotationDocument9 pagesExperiment 5 Properties of Carbohydrates: Solubility, Reactivity, and Specific RotationCiara marie BernardoNo ratings yet
- Natural PolymersDocument59 pagesNatural PolymersOlayanjuNo ratings yet
- Biochemistry of Glycolysis and Metabolism EnzymesDocument48 pagesBiochemistry of Glycolysis and Metabolism EnzymesBrother GeorgeNo ratings yet
- 11 GlycolysisDocument51 pages11 GlycolysisGianna Kristen MirandaNo ratings yet
- CarbohydratesDocument27 pagesCarbohydrateseugieniogienioNo ratings yet
- Exm N X11 Chem Biomol ADocument28 pagesExm N X11 Chem Biomol Asumair hejibNo ratings yet
- Kraft Reactin-Dye SynthesisDocument7 pagesKraft Reactin-Dye Synthesisgulft1777No ratings yet
- محاضره ٤Document25 pagesمحاضره ٤ecosystem200410myNo ratings yet
- 05-Minggu V Dan VI - BiokimiaDocument91 pages05-Minggu V Dan VI - Biokimiahanny nuguNo ratings yet
- CarbohydratesDocument28 pagesCarbohydratesMae Codium GallentesNo ratings yet
- BIOC 200 Lec # 5 - LIPIDSDocument44 pagesBIOC 200 Lec # 5 - LIPIDSEslam NassarNo ratings yet
- 7.014 Handout: Biochemistry I-IIIDocument8 pages7.014 Handout: Biochemistry I-IIIRiriiiNo ratings yet
- Dr.H.Mohammad Hanafi, Mbbs (Syd.) .MS.: Fakultas Kedokteran UnairDocument48 pagesDr.H.Mohammad Hanafi, Mbbs (Syd.) .MS.: Fakultas Kedokteran UnairahmustofaNo ratings yet
- Carbohydrates 1: MSL: BiochemistryDocument50 pagesCarbohydrates 1: MSL: Biochemistryنجوي عبدالوهابNo ratings yet
- 1 ChoDocument79 pages1 ChoFrance Jan First SaplacoNo ratings yet
- Fatty Acid Oxidation: Molecular Biochemistry IIDocument39 pagesFatty Acid Oxidation: Molecular Biochemistry IIDozdi100% (1)
- Carbohydrates: Molecular BiochemistryDocument36 pagesCarbohydrates: Molecular BiochemistryDozdiNo ratings yet
- Lipids 2Document6 pagesLipids 2cumbredinNo ratings yet
- Biokimia1 CarbohydratesDocument52 pagesBiokimia1 CarbohydratesfinaNo ratings yet
- Overview of Alcohol Production 07042020Document66 pagesOverview of Alcohol Production 07042020PujitNo ratings yet
- Glycolysis Glikolisis - Serangkaian Reaksi Katabolisme Glukosa Untuk Menghasilkan Energi (ATP) - Pemecahan Glukosa Menjadi Asam PiruvatDocument40 pagesGlycolysis Glikolisis - Serangkaian Reaksi Katabolisme Glukosa Untuk Menghasilkan Energi (ATP) - Pemecahan Glukosa Menjadi Asam PiruvatWidri LianahNo ratings yet
- MC2 Lec11 Catabolic PathwaysDocument61 pagesMC2 Lec11 Catabolic PathwaysLauren CarlosIINo ratings yet
- Drawing Haworth ProjectionsDocument6 pagesDrawing Haworth ProjectionschoconoodlesNo ratings yet
- Carbohydrate MetabolismDocument52 pagesCarbohydrate MetabolismLucy ZuluNo ratings yet
- Pathways of Glucose Metabolism: Blood Glucose Rna & DnaDocument17 pagesPathways of Glucose Metabolism: Blood Glucose Rna & DnaReem AkraNo ratings yet
- Osteoprorosis Therapy Management, Beyond Safety and Efficacy (Once Yearly Treatment)Document29 pagesOsteoprorosis Therapy Management, Beyond Safety and Efficacy (Once Yearly Treatment)ian ismeNo ratings yet
- Pathways of Glucose Metabolism: Blood Glucose Rna & DnaDocument17 pagesPathways of Glucose Metabolism: Blood Glucose Rna & DnaAhmedNo ratings yet
- Biomolecules and Polymers-02 - Solved ProblemsDocument11 pagesBiomolecules and Polymers-02 - Solved ProblemsRaju SinghNo ratings yet
- Shikimic PathwayDocument27 pagesShikimic Pathwayherukassei100% (1)
- Buletine DigestivDocument5 pagesBuletine DigestivStanescuRozicaNo ratings yet
- YuvhDocument1 pageYuvhStanescuRozicaNo ratings yet
- Yotn0rweupf0r VB 34Document1 pageYotn0rweupf0r VB 34StanescuRozicaNo ratings yet
- Rby56 Buy6 12Document1 pageRby56 Buy6 12StanescuRozicaNo ratings yet
- Etyousti f12Document1 pageEtyousti f12StanescuRozicaNo ratings yet
- FGHJK 7 Tbyh 45Document1 pageFGHJK 7 Tbyh 45StanescuRozicaNo ratings yet
- Hgfds ReDocument1 pageHgfds ReStanescuRozicaNo ratings yet
- Farmacologie in ComprimateDocument414 pagesFarmacologie in ComprimateEliza Iancu100% (14)
- Nbh9trbucmqwc 43Document1 pageNbh9trbucmqwc 43StanescuRozicaNo ratings yet
- Nr6inbeytrvbnio P.docx ReferDocument1 pageNr6inbeytrvbnio P.docx ReferStanescuRozicaNo ratings yet
- Pmgcrwpfeior Otpugiro STR - Docx BGDocument1 pagePmgcrwpfeior Otpugiro STR - Docx BGStanescuRozicaNo ratings yet
- Presentation 1Document1 pagePresentation 1StanescuRozicaNo ratings yet
- VJNG Ghjfgujrv Reutjf Frevghjfrpi FFDV F VGH Bru G Gvrfmccuv GFDSV Gujssadam HGF Uhg Vuhgtujgvsearfvghtgjfddighjvsrerivurvgjfds Ugvitr Stggrewpaosrkfgnmf IrfDocument1 pageVJNG Ghjfgujrv Reutjf Frevghjfrpi FFDV F VGH Bru G Gvrfmccuv GFDSV Gujssadam HGF Uhg Vuhgtujgvsearfvghtgjfddighjvsrerivurvgjfds Ugvitr Stggrewpaosrkfgnmf IrfStanescuRozicaNo ratings yet
- Nr6inbeytrvbnio P.docx Refer - Docx 3333333Document1 pageNr6inbeytrvbnio P.docx Refer - Docx 3333333StanescuRozicaNo ratings yet
- Staying fit with exercise tipsDocument1 pageStaying fit with exercise tipsStanescuRozicaNo ratings yet
- Basic Integration Problems #1Document4 pagesBasic Integration Problems #1John Marlo GorobaoNo ratings yet
- 1964 Plymouth Barracuda Landau (Factory Prototype W/ Targa Top) - Photos & CorrespondenceDocument12 pages1964 Plymouth Barracuda Landau (Factory Prototype W/ Targa Top) - Photos & CorrespondenceJoseph GarzaNo ratings yet
- Nexus Capital Management Annual ReportDocument7 pagesNexus Capital Management Annual ReportDavid VasquezNo ratings yet
- TR019999012181 enDocument37 pagesTR019999012181 enAliTronic1972No ratings yet
- Hydrocarbon ReactionsDocument2 pagesHydrocarbon ReactionsJessa Libo-onNo ratings yet
- Kti Semuanya-DikonversiDocument59 pagesKti Semuanya-DikonversigacikNo ratings yet
- 2 Phantom Forces Gui Scripts in OneDocument11 pages2 Phantom Forces Gui Scripts in OneAthallah Rafif JNo ratings yet
- Troubleshooting Directory for LHB Type RMPU EOG AC CoachesDocument44 pagesTroubleshooting Directory for LHB Type RMPU EOG AC Coachesdivisional electrical engg100% (6)
- DL Throughput Improvement with Aperiodic CQI ReportingDocument10 pagesDL Throughput Improvement with Aperiodic CQI ReportingAqeel HasanNo ratings yet
- Man and Mystery Vol 13 - Monsters and Cryptids (Rev06)Document139 pagesMan and Mystery Vol 13 - Monsters and Cryptids (Rev06)Pablo Jr AgsaludNo ratings yet
- 12.3 Operation Qualification Protocol For Laminar Air Flow UnitDocument4 pages12.3 Operation Qualification Protocol For Laminar Air Flow UnituzairNo ratings yet
- VTBS 20-3DDocument1 pageVTBS 20-3Dwong keen faivNo ratings yet
- DD 24 DD 22Document2 pagesDD 24 DD 22evega97No ratings yet
- Pol Science ProjectDocument18 pagesPol Science ProjectAnshu SharmaNo ratings yet
- Referencia 8Document130 pagesReferencia 8Kimiko SullonNo ratings yet
- TN 015 Overrides 15Document8 pagesTN 015 Overrides 15Luis SantosNo ratings yet
- OK - EndUserGuideHoseSafetyInstituteDocument8 pagesOK - EndUserGuideHoseSafetyInstituteSunil GhosalkarNo ratings yet
- Iso 4309 2017Document10 pagesIso 4309 2017C. de JongNo ratings yet
- Pulsar220S PLANOS PDFDocument32 pagesPulsar220S PLANOS PDFJuan Jose MoralesNo ratings yet
- Fe412farm Module N Cost BenefitDocument34 pagesFe412farm Module N Cost BenefitYaswanth NaikNo ratings yet
- SEO-Optimized Title for Quantitative Techniques for Business-II Exam DocumentDocument4 pagesSEO-Optimized Title for Quantitative Techniques for Business-II Exam DocumentEthan WillsNo ratings yet
- AEIOU Framework - Case Study On Agriculture Domain (Automatic Drip Tube Irrigation System)Document30 pagesAEIOU Framework - Case Study On Agriculture Domain (Automatic Drip Tube Irrigation System)Kunal TalegaonkarNo ratings yet
- SG PB Imds1350Document103 pagesSG PB Imds1350zky0108No ratings yet
- Ship Construction IonsDocument25 pagesShip Construction Ionsanon-49038083% (6)
- Sui Gas Connection FormsDocument1 pageSui Gas Connection Formsare50% (2)
- 3D Bioprinting From The Micrometer To Millimete 2017 Current Opinion in BiomDocument7 pages3D Bioprinting From The Micrometer To Millimete 2017 Current Opinion in Biomrrm77No ratings yet
- Taoism and The KabbalahDocument26 pagesTaoism and The KabbalahJim Weaver100% (2)
- Well Components: Petroleum Production Engineering. DOI: © 2007 Elsevier Inc. All Rights ReservedDocument15 pagesWell Components: Petroleum Production Engineering. DOI: © 2007 Elsevier Inc. All Rights Reservedsoran najebNo ratings yet
- Radiation Safety Procedure Rev.08Document41 pagesRadiation Safety Procedure Rev.08manz29No ratings yet
- D.K.Pandey: Lecture 1: Growth and Decay of Current in RL CircuitDocument5 pagesD.K.Pandey: Lecture 1: Growth and Decay of Current in RL CircuitBBA UniversityNo ratings yet