You might also like
- Endocrine System: A Tutorial Study GuideFrom EverandEndocrine System: A Tutorial Study GuideRating: 5 out of 5 stars5/5 (1)
- Endocrine System Endocrine System: "The Body Regulator"Document31 pagesEndocrine System Endocrine System: "The Body Regulator"angelus008No ratings yet
- Receptor Mechanisms: - Receptor Are Proteins With High Affinity To ADocument45 pagesReceptor Mechanisms: - Receptor Are Proteins With High Affinity To AbevorsiNo ratings yet
- Test Bank For Guyton and Hall Textbook of Medical Physiology 13th Edition by John e Hall 005 2Document10 pagesTest Bank For Guyton and Hall Textbook of Medical Physiology 13th Edition by John e Hall 005 2courtneyriveraefjpadcmyx100% (21)
- Smooth and Cardiac MusclesDocument55 pagesSmooth and Cardiac MusclesIni UsenNo ratings yet
- A&P Final Exam Review Study GuideDocument21 pagesA&P Final Exam Review Study GuideLisa HuttonNo ratings yet
- Immune Response DRPath Part 1Document71 pagesImmune Response DRPath Part 1SITI NUR FA'IZULBA'EDAH BINTI MAT RAZINo ratings yet
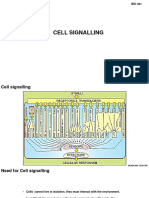
- Cell SignallingDocument20 pagesCell SignallingyanaNo ratings yet
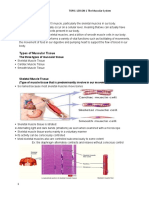
- Anaphy Muscular SystemDocument54 pagesAnaphy Muscular SystemZosia SageNo ratings yet
- Gastro-Intestinal Hormones & TransmittersDocument23 pagesGastro-Intestinal Hormones & TransmittersAlbert ChuwaNo ratings yet
- 397 - Musculoskeletal Physiology) Smooth MuscleDocument5 pages397 - Musculoskeletal Physiology) Smooth Musclevkpremiyahoo.comNo ratings yet
- Cell SignalingDocument53 pagesCell SignalingKaushiki KalravNo ratings yet
- Cell Signaling - ImpDocument11 pagesCell Signaling - Impkrishnarajagopal2011No ratings yet
- Endocrine Physiology: Dale Buchanan Hales, PHD Department of Physiology & BiophysicsDocument64 pagesEndocrine Physiology: Dale Buchanan Hales, PHD Department of Physiology & Biophysicshamako02No ratings yet
- Cell SignalingDocument43 pagesCell SignalingRIFAM zNo ratings yet
- Signal and Signal Transduction For Mbs1 2014Document52 pagesSignal and Signal Transduction For Mbs1 2014Jefri SandikaNo ratings yet
- Cell Communications and Signal Transduction: Biokimia Dan Biologi Molekuler FK UntarDocument112 pagesCell Communications and Signal Transduction: Biokimia Dan Biologi Molekuler FK UntarAlbert TandyNo ratings yet
- Chapter 9Document16 pagesChapter 9skywalkerNo ratings yet
- Endocrine System PDFDocument38 pagesEndocrine System PDFUlfat NiazyNo ratings yet
- K7 - Komunikasi Antar SelDocument30 pagesK7 - Komunikasi Antar SelJuliana Sari HarahapNo ratings yet
- Molecular Endocrine, 1Document50 pagesMolecular Endocrine, 1Dedi IrwansyahNo ratings yet
- Cell Signaling: Your Ad Could Be Here! Click Here To InquireDocument7 pagesCell Signaling: Your Ad Could Be Here! Click Here To InquireRionNo ratings yet
- Chapter 15- Jnu Cellular Signaling - 複本Document49 pagesChapter 15- Jnu Cellular Signaling - 複本Wai Kwong ChiuNo ratings yet
- Review of The Endocrine System: Dr. Amiruddin Eso, M.Kes Department of Physiology Medical Faculty Halu Oleo UniversityDocument75 pagesReview of The Endocrine System: Dr. Amiruddin Eso, M.Kes Department of Physiology Medical Faculty Halu Oleo UniversityWidodo MuhammadNo ratings yet
- UG SEM-6 (H) - CC-XIII Signal TransductionDocument22 pagesUG SEM-6 (H) - CC-XIII Signal Transductionn7sspyywc6No ratings yet
- Summary of Sabas Notes (Lab 3)Document21 pagesSummary of Sabas Notes (Lab 3)anzalnaamanNo ratings yet
- Cell Communication Class Notes Presentation Topics 4.1 To 4.2 2020Document48 pagesCell Communication Class Notes Presentation Topics 4.1 To 4.2 2020Lavinia DonaldNo ratings yet
- 1st Lec On Endo Physiology by DR RoomiDocument24 pages1st Lec On Endo Physiology by DR RoomiMudassar RoomiNo ratings yet
- Endocrine SystemDocument34 pagesEndocrine SystemNada DardeerNo ratings yet
- Cell CommunicationDocument24 pagesCell Communicationanbuselvanselvan59No ratings yet
- Cell Bio 5 Cell Communication 2021 PDFDocument31 pagesCell Bio 5 Cell Communication 2021 PDFaboodmr8nblahNo ratings yet
- Cell Signalling and CommunicationsDocument4 pagesCell Signalling and CommunicationsJames IbrahimNo ratings yet
- Lect. 10Document27 pagesLect. 10أحمدشريفNo ratings yet
- Biocehem AssignDocument47 pagesBiocehem Assignwendemeneh abateNo ratings yet
- Hormonal Signal Transduction TutorialDocument45 pagesHormonal Signal Transduction TutorialnaghalfatihNo ratings yet
- Intercellular Communication: Cell and Molecular PhysiologyDocument30 pagesIntercellular Communication: Cell and Molecular PhysiologyO'Brien AlfredNo ratings yet
- Cell SignalingDocument20 pagesCell Signalingnarehan13No ratings yet
- Cell Signaling Write UpDocument4 pagesCell Signaling Write Upinfinity chronusNo ratings yet
- Cell Signalling: Prof. Dr. Selma YILMAZER Medical Biology DepartmentDocument46 pagesCell Signalling: Prof. Dr. Selma YILMAZER Medical Biology DepartmentNasan ShehadaNo ratings yet
- Mecanism of Action of Hormones 9.6.20Document46 pagesMecanism of Action of Hormones 9.6.20Mohammad ZayanNo ratings yet
- Overview of Cell Signal Transduction 101Document13 pagesOverview of Cell Signal Transduction 101Ilac Tristan BernardoNo ratings yet
- ENDOCRIOLOGYDocument138 pagesENDOCRIOLOGYAqurate BuildersNo ratings yet
- Cell Signalling BasicsDocument26 pagesCell Signalling Basics6ypywxdqmkNo ratings yet
- Mechanism of Hormone ActionDocument2 pagesMechanism of Hormone Actionegfr3yfgNo ratings yet
- Cells Typically Communicate Using Chemical SignalsDocument5 pagesCells Typically Communicate Using Chemical SignalsrmndzrmeNo ratings yet
- DR Norhan PDF Lecture 3 Signal Transduction For Students Elalamein 2021-22Document4 pagesDR Norhan PDF Lecture 3 Signal Transduction For Students Elalamein 2021-22Ahmed MagdyNo ratings yet
- Hormone FinalDocument36 pagesHormone FinalRajkishor YadavNo ratings yet
- CytokinesDocument29 pagesCytokinesAhmed AlosaiqueNo ratings yet
- 5endocrlect3 191011165740Document55 pages5endocrlect3 191011165740OROKE JOHN EJENo ratings yet
- Plant Cell SignalingDocument35 pagesPlant Cell SignalingAmelia Sukma Wati325No ratings yet
- Cell Communication NotesDocument3 pagesCell Communication NoteselyssaNo ratings yet
- Biochemistry - Signal TransductionDocument4 pagesBiochemistry - Signal Transductionsebasmarq2001No ratings yet
- Signalling in CellsDocument11 pagesSignalling in CellsAditi JhaNo ratings yet
- 1 The Progress of Cell SignalingDocument72 pages1 The Progress of Cell SignalingVincent LangatNo ratings yet
- Lecture 9 Part 1Document33 pagesLecture 9 Part 1Kimberly GrahamNo ratings yet
- The Endocrine System: HormonesDocument948 pagesThe Endocrine System: HormonesMahaaaNo ratings yet
- Cell Signaling-ClassDocument49 pagesCell Signaling-ClassDeepanshu rawatNo ratings yet
- Biochemistry Assignment Submitted To: Dr. Noureen Submitted byDocument15 pagesBiochemistry Assignment Submitted To: Dr. Noureen Submitted bySalman KhanNo ratings yet
- Cell Signaling-2021Document16 pagesCell Signaling-2021VINE TUBENo ratings yet
- Endocrine System: University of SurreyDocument13 pagesEndocrine System: University of SurreyqhqhqNo ratings yet
- Cell Signaling PathwaysDocument18 pagesCell Signaling PathwaysJean Marcel Sousa LiraNo ratings yet
- Ch. 6: Communication, Integration & HomeostasisDocument31 pagesCh. 6: Communication, Integration & HomeostasisromioalarabNo ratings yet
- E N D o C R I N o L o G Y: Dr. Victoria G. Giango Chair, Dept. of PhysiologyDocument23 pagesE N D o C R I N o L o G Y: Dr. Victoria G. Giango Chair, Dept. of Physiologyanon-271548No ratings yet
- Regulation of Gene Expression in Eukaryotes (Light)Document46 pagesRegulation of Gene Expression in Eukaryotes (Light)Amit PaulNo ratings yet
- Adrenoceptor DrugsDocument30 pagesAdrenoceptor DrugsLloyd Selorio ArboledaNo ratings yet
- Muscular TissueDocument65 pagesMuscular TissueMitzel SapaloNo ratings yet
- Microscopic Anatomy of A Skeletal Muscle FiberDocument33 pagesMicroscopic Anatomy of A Skeletal Muscle FiberKumar MSNo ratings yet
- Thesis Michiel Maarten SteinzDocument79 pagesThesis Michiel Maarten SteinzShukr Wesman BlbasNo ratings yet
- Group VII - Smooth Muscle ContractionDocument7 pagesGroup VII - Smooth Muscle ContractioniqbalNo ratings yet
- 5-Heart FailureDocument12 pages5-Heart FailureJericho De GuzmanNo ratings yet
- Wearing On Her Nerves Case Study - Werner Williams Mccool Floyd AhmadDocument50 pagesWearing On Her Nerves Case Study - Werner Williams Mccool Floyd Ahmadapi-320365850100% (2)
- Physiology. Hrd.Document549 pagesPhysiology. Hrd.Sʌɩĸʌt PʌʋɭNo ratings yet
- Sarcoplasmic Hypertrophy in Skeletal Muscle A ScieDocument16 pagesSarcoplasmic Hypertrophy in Skeletal Muscle A ScieRosaneLacerdaNo ratings yet
- Skeletal Muscle: A Brief Review of Structure and Function: Calcified Tissue International October 2014Document14 pagesSkeletal Muscle: A Brief Review of Structure and Function: Calcified Tissue International October 2014Anonymous K9tmd7T0PzNo ratings yet
- 4.1.1 Draw and Label A Diagram of A Motor Unit.: Movement AnalysisDocument30 pages4.1.1 Draw and Label A Diagram of A Motor Unit.: Movement AnalysisChristyle AquinoNo ratings yet
- Airway Smooth Muscle RelaxationDocument8 pagesAirway Smooth Muscle Relaxationmirza_baig_46No ratings yet
- Chapter Six The Muscular SystemDocument30 pagesChapter Six The Muscular SystemabiyotNo ratings yet
- Pathophysiology On ArrhythmiaDocument34 pagesPathophysiology On ArrhythmiaYhr Yh100% (1)
- SN5 Mark Scheme Sum 2007Document10 pagesSN5 Mark Scheme Sum 2007EmmaNo ratings yet
- 3 - Cardiac Contractility - Ass. Prof. Doaa Abou-Bakr - 2020Document8 pages3 - Cardiac Contractility - Ass. Prof. Doaa Abou-Bakr - 2020Hossam BaniisNo ratings yet
- Phs 201 For Nursing & Mls StudentsDocument104 pagesPhs 201 For Nursing & Mls StudentsayotomiwatemisanNo ratings yet
- Guyton Hall PHYSIOLOGY Chapter 8 PDFDocument7 pagesGuyton Hall PHYSIOLOGY Chapter 8 PDFOsman NazirNo ratings yet
- Biology Portion of Etea Medical Test 2019Document21 pagesBiology Portion of Etea Medical Test 2019Ihtisham Ul Haq75% (4)
- Creatine and Caffeine - ConsiderationsDocument17 pagesCreatine and Caffeine - ConsiderationsVanessa BarrosNo ratings yet
- Sliding Filament TheoryDocument9 pagesSliding Filament Theoryellandhae100% (1)
- Biochem Report Muscle and CytoskeletonDocument87 pagesBiochem Report Muscle and CytoskeletonKate Lynne CamonayanNo ratings yet
- The Enzyme List Class 7 - Translocases: ExplorenzDocument35 pagesThe Enzyme List Class 7 - Translocases: ExplorenzgustasconNo ratings yet
- Creatinin and CaffeineDocument33 pagesCreatinin and CaffeineNina RianaNo ratings yet