You might also like
- Plasticity FascialDocument13 pagesPlasticity FascialMichael DuranNo ratings yet
- The Fascia The Forgotten Structure PDFDocument12 pagesThe Fascia The Forgotten Structure PDFJorge100% (1)
- Bio Dynamics The Fascia of The Pelvic Floor Model of TensegrityDocument8 pagesBio Dynamics The Fascia of The Pelvic Floor Model of TensegritygabrielsantosmsNo ratings yet
- Fascia Forgotten StructureDocument12 pagesFascia Forgotten StructureFraan GutierrezNo ratings yet
- The Neutral Spine PrincipleDocument12 pagesThe Neutral Spine Principlesabrina100% (1)
- M. Rowane - Core Concepts in Osteopathic Manipulative MedicineDocument55 pagesM. Rowane - Core Concepts in Osteopathic Manipulative Medicinedruidsamsung100% (1)
- CLARENCE TIU - Criminal Law 1 Notes (Last Edit-Jan2018) PDFDocument163 pagesCLARENCE TIU - Criminal Law 1 Notes (Last Edit-Jan2018) PDFRica Dominguez100% (3)
- What Is Functional NeurologyDocument2 pagesWhat Is Functional NeurologydrtimadamsNo ratings yet
- Biological Effects of Direct and Indirect Manipulation of The Fascial System - Parravicini2017Document11 pagesBiological Effects of Direct and Indirect Manipulation of The Fascial System - Parravicini2017Angel AngelNo ratings yet
- Lossing DiagnosingTreatingDu PDFDocument33 pagesLossing DiagnosingTreatingDu PDFDiana SchlittlerNo ratings yet
- Chest DrainageDocument22 pagesChest DrainageAmeliaM100% (7)
- Fascial Plasticity A New Neurobiological Explanation Schleip Parte 1Document9 pagesFascial Plasticity A New Neurobiological Explanation Schleip Parte 1Lucia MerlinoNo ratings yet
- Reflex ArcDocument16 pagesReflex ArcNiaz Ahammed50% (2)
- Workshop 1 PowerpointDocument10 pagesWorkshop 1 PowerpointCarlos CorreaNo ratings yet
- The Bioenergetic Model in Osteopathic Diagnosis and Treatment: An FAAO Thesis, Part 1Document9 pagesThe Bioenergetic Model in Osteopathic Diagnosis and Treatment: An FAAO Thesis, Part 1nish0420100% (1)
- The Fascia As Organ of Innerness 2015 EN ChapterDocument16 pagesThe Fascia As Organ of Innerness 2015 EN ChapterJorge Flores TorresNo ratings yet
- Mechanobiology and Diseases of Mechanotransduction by IngberDocument14 pagesMechanobiology and Diseases of Mechanotransduction by IngberTensegrity Wiki100% (1)
- The Signficance of RespirationDocument12 pagesThe Signficance of RespirationApril Dawn100% (1)
- Human KinesiologyDocument26 pagesHuman KinesiologyTrinidad Farías PiñeiroNo ratings yet
- 2010 The Fascial Manipulation Technique and Its Biomechanical Model: A Guide To The Human Fascial SystemDocument3 pages2010 The Fascial Manipulation Technique and Its Biomechanical Model: A Guide To The Human Fascial System1Rogerio100% (1)
- Sensorimotor Integration For FunctionalDocument17 pagesSensorimotor Integration For FunctionalFuncionalidad DiversaNo ratings yet
- The Pain and Movement Reasoning ModelDocument7 pagesThe Pain and Movement Reasoning ModelRamiro Avendaño Rebolledo100% (1)
- Osteopathic Fascial ManipulationDocument9 pagesOsteopathic Fascial ManipulationnunoNo ratings yet
- Cranial Rhythmic Impulse Related To The Traube-Hering-Mayer OscillationDocument11 pagesCranial Rhythmic Impulse Related To The Traube-Hering-Mayer Oscillationthe_walnutNo ratings yet
- Old Ideas: The Specificity Theory of PainDocument8 pagesOld Ideas: The Specificity Theory of Paingeloss23150% (2)
- OMDTPDocument625 pagesOMDTPromi_brener100% (1)
- AAO FDM Indroduction 1994 PDFDocument12 pagesAAO FDM Indroduction 1994 PDFHONGJYNo ratings yet
- From Clinical Experience To A Model For The Human Fascial SystemDocument9 pagesFrom Clinical Experience To A Model For The Human Fascial SystemMohamed Magdy ElMeligieNo ratings yet
- OPP Course Review: John Garlitz, DO Deborah Schmidt, DO Opp 2 April 15, 2015 8:10 AMDocument248 pagesOPP Course Review: John Garlitz, DO Deborah Schmidt, DO Opp 2 April 15, 2015 8:10 AMmina botross100% (1)
- 3.5 Mundane MaterialsDocument140 pages3.5 Mundane MaterialsJames LewisNo ratings yet
- Chapter 5 Human Movement ScienceDocument30 pagesChapter 5 Human Movement ScienceVu Van Cuong100% (1)
- FasciaDocument12 pagesFasciaRodolfo CiqueleiroNo ratings yet
- Pain and NeuromatrixDocument5 pagesPain and NeuromatrixBharathGopinathNo ratings yet
- Ingber Lecture: From Cellular Mechanotransduction To Biologically Inspired EngineeringDocument14 pagesIngber Lecture: From Cellular Mechanotransduction To Biologically Inspired EngineeringTensegrity Wiki100% (2)
- Chapmans Reflex 479Document5 pagesChapmans Reflex 479Huram-abi0% (1)
- Phantom Limbs and Plastic BrainDocument19 pagesPhantom Limbs and Plastic BrainZinneRah RahManNo ratings yet
- Jim Jealous, - Healing and The Natural WorldDocument8 pagesJim Jealous, - Healing and The Natural WorldSeda Joseph Saine100% (1)
- Motifs: The Heart of Listening Volume One by Hugh MilneDocument1 pageMotifs: The Heart of Listening Volume One by Hugh MilneSimon FalconiNo ratings yet
- The Five Diaphragms in Osteopathic Manipulative MeDocument14 pagesThe Five Diaphragms in Osteopathic Manipulative MeShaun Tyler100% (1)
- Posture, Dynamic Stability, and Voluntary MovementDocument18 pagesPosture, Dynamic Stability, and Voluntary MovementAnonymous IXswcnWNo ratings yet
- The Organ of Form: Towards A Theory of Biological ShapeDocument11 pagesThe Organ of Form: Towards A Theory of Biological Shapesusanne voraNo ratings yet
- The Masculoskeletal System & Tissue MechanicsDocument29 pagesThe Masculoskeletal System & Tissue MechanicsEdara Rajesh KumarNo ratings yet
- LTC 4240 Art Integration Lesson Plan TemplateDocument8 pagesLTC 4240 Art Integration Lesson Plan Templateapi-245789430No ratings yet
- Clinical Neurodynamics and Sports Medicine: Origins and DevelopmentDocument8 pagesClinical Neurodynamics and Sports Medicine: Origins and DevelopmentWahid NasrudinNo ratings yet
- Cranial Osteopathy Its Fate Seems ClearDocument3 pagesCranial Osteopathy Its Fate Seems ClearInvincy JYNo ratings yet
- Futurist CinemaDocument31 pagesFuturist CinemaguidogiglioNo ratings yet
- Philosophy and Education in MalaysiaDocument42 pagesPhilosophy and Education in MalaysiaThiyya SylviaNo ratings yet
- Effectiveness of Fascial Manipulation Method FMRDocument9 pagesEffectiveness of Fascial Manipulation Method FMRDr Robin Sharma SPINE ACTIONNo ratings yet
- Transverse Rotation of The Segments of The Lower Extremity in LocomotionDocument15 pagesTransverse Rotation of The Segments of The Lower Extremity in Locomotionpetcudaniel100% (1)
- CV4 EfectDocument21 pagesCV4 EfectNicole HermosillaNo ratings yet
- Revealing The Hidden: The Epiphanic Dimension of Games and SportDocument31 pagesRevealing The Hidden: The Epiphanic Dimension of Games and SportXimena BantarNo ratings yet
- Facilitated SegmentsDocument6 pagesFacilitated Segmentsismaelvara6568No ratings yet
- Embryogenesis SheaDocument4 pagesEmbryogenesis Sheaimageorge13100% (1)
- The Fascial Distortion Model BookDocument10 pagesThe Fascial Distortion Model BooksimonyanNo ratings yet
- The Helical Ventricular Myocardial Band of Torrent-GuaspDocument5 pagesThe Helical Ventricular Myocardial Band of Torrent-GuaspTeaKovacevicNo ratings yet
- ConstipationDocument22 pagesConstipationash ash100% (1)
- Pes Cavus and Hereditary Neuropathies When A Relationship Should Be SuspectedDocument5 pagesPes Cavus and Hereditary Neuropathies When A Relationship Should Be SuspectedSuhar Tomi100% (1)
- How To Take Your TimeDocument3 pagesHow To Take Your TimePhương Hân TrầnNo ratings yet
- BiomechanicsDocument17 pagesBiomechanicss_chandrubioNo ratings yet
- Pain Book FeinDocument102 pagesPain Book FeinHansen NielsNo ratings yet
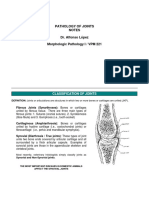
- Pathology of JointsDocument13 pagesPathology of JointswobblegobbleNo ratings yet
- Surgical Technique For Combined D Wyer Calcaneal Osteotomy and PeronealDocument6 pagesSurgical Technique For Combined D Wyer Calcaneal Osteotomy and PeronealleahbayNo ratings yet
- Membranes, Mitochondria, and Connective TissuesFrom EverandMembranes, Mitochondria, and Connective TissuesIsidore GershNo ratings yet
- Tensegrity Beam Wave Dynamics 2019Document20 pagesTensegrity Beam Wave Dynamics 2019Tensegrity WikiNo ratings yet
- Pinuad Dynamics CRCDocument62 pagesPinuad Dynamics CRCTensegrity WikiNo ratings yet
- Marching Procedure For Tensegrity 2007Document26 pagesMarching Procedure For Tensegrity 2007Tensegrity WikiNo ratings yet
- Han LMI Struct Linear Cdc03Document6 pagesHan LMI Struct Linear Cdc03Tensegrity WikiNo ratings yet
- Weiwei FSN CDC42Document6 pagesWeiwei FSN CDC42Tensegrity WikiNo ratings yet
- Optimal Topology and Geometry For Controllable Tensegrity Systems by Jager, Masic, SkeltonDocument6 pagesOptimal Topology and Geometry For Controllable Tensegrity Systems by Jager, Masic, SkeltonTensegrity WikiNo ratings yet
- Equilibrium Conditions of A Tensegrity Structure by Williamson, Skelton, Han 2002Document18 pagesEquilibrium Conditions of A Tensegrity Structure by Williamson, Skelton, Han 2002Tensegrity WikiNo ratings yet
- Structure Optimization Approach For Minimum-Time Reconfiguration of Tensegrity Systems by Aldrich, SkeltonDocument12 pagesStructure Optimization Approach For Minimum-Time Reconfiguration of Tensegrity Systems by Aldrich, SkeltonTensegrity WikiNo ratings yet
- Stiffness of Planar Tensegrity Truss Topologies by Jager, Skelton 2006Document23 pagesStiffness of Planar Tensegrity Truss Topologies by Jager, Skelton 2006Tensegrity WikiNo ratings yet
- Intro To Mechanics of Tensegrity StructuresDocument6 pagesIntro To Mechanics of Tensegrity StructuresTensegrity WikiNo ratings yet
- Tensegrity Airfoils Henrickson, Skelton, Valasek 2016Document25 pagesTensegrity Airfoils Henrickson, Skelton, Valasek 2016Tensegrity WikiNo ratings yet
- Deployment of A Class 2 Tensegrity Boom by Pinauda, Solaria, SkeltonDocument8 pagesDeployment of A Class 2 Tensegrity Boom by Pinauda, Solaria, SkeltonTensegrity WikiNo ratings yet
- Andy WarholDocument4 pagesAndy WarholAngel Angeleri-priftis.No ratings yet
- B.Sc. Microbiology and Clinical Lab TechnologyDocument80 pagesB.Sc. Microbiology and Clinical Lab TechnologyDawnNo ratings yet
- Magic City Acceptance Center ApplicationDocument55 pagesMagic City Acceptance Center ApplicationJonece DuniganNo ratings yet
- Thesis Manual 2008Document138 pagesThesis Manual 2008Afri Kesling Document'sNo ratings yet
- Final Spl-Enriquez - Ra 9165Document40 pagesFinal Spl-Enriquez - Ra 9165EphraimEnriquezNo ratings yet
- Lat235 Pset 8Document5 pagesLat235 Pset 8Muhamad Arthura Arya FarrozNo ratings yet
- Impact of Emotional Intelligence On Work Life BalanceDocument17 pagesImpact of Emotional Intelligence On Work Life BalanceAakshi DNo ratings yet
- CGI Programming in CDocument27 pagesCGI Programming in CPiya BaradNo ratings yet
- The Four Confidences EbookDocument38 pagesThe Four Confidences EbookIgor PopadicNo ratings yet
- Saint Paul University Surigao College of Business and Technology Grading SystemDocument9 pagesSaint Paul University Surigao College of Business and Technology Grading SystemKeziah AliwanagNo ratings yet
- Microlife BP 3BT0 A (2) ManualDocument50 pagesMicrolife BP 3BT0 A (2) ManualVanessa MarinNo ratings yet
- Cost Acctg FohDocument37 pagesCost Acctg FohRizza Christine Thereza UsbalNo ratings yet
- Clone Graph - LeetCodeDocument1 pageClone Graph - LeetCodejon monroeNo ratings yet
- Social Change and EvolutionDocument15 pagesSocial Change and Evolutioncooooool1927100% (2)
- Engr1513 Syllabus FA20 AbetDocument3 pagesEngr1513 Syllabus FA20 AbetAustin MitchellNo ratings yet
- 13 PaladiniDocument9 pages13 PaladinialdybebikaNo ratings yet
- Morkunas Và Rudiene, 2020. The - Impact - of - Social - Servicescape - Factors - On - CustoDocument15 pagesMorkunas Và Rudiene, 2020. The - Impact - of - Social - Servicescape - Factors - On - CustoHương GiangNo ratings yet
- NavrasDocument2 pagesNavrasAayushi PandeyNo ratings yet
- AG LetterDocument2 pagesAG Lettermylene sangriaNo ratings yet
- Reporting ScripttDocument4 pagesReporting Scripttnicole besarioNo ratings yet
- The Bom 100 Day ChallengeDocument4 pagesThe Bom 100 Day Challengeapi-336153536No ratings yet
- Bambara LanguageDocument6 pagesBambara LanguageAndré DeFranciaNo ratings yet
- Medieval Project and RubricDocument2 pagesMedieval Project and RubricZun Phyo Thiri OoNo ratings yet
- Scheme of Work 2010Document22 pagesScheme of Work 2010Sivabalan BionicNo ratings yet