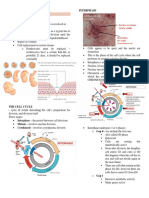
You might also like
- Carotenoids Other Than Vitamin A — III: Third International Symposium on Carotenoids Other Than Vitamin AFrom EverandCarotenoids Other Than Vitamin A — III: Third International Symposium on Carotenoids Other Than Vitamin ANo ratings yet
- Betalain Production Is Possible in Anthocyanin-Producing Plant Species Given The Presence of DOPA-dioxygenase and L-DOPADocument13 pagesBetalain Production Is Possible in Anthocyanin-Producing Plant Species Given The Presence of DOPA-dioxygenase and L-DOPARosane LiégeNo ratings yet
- Carotenoid Metabolism in Plants: Molecular PlantDocument15 pagesCarotenoid Metabolism in Plants: Molecular PlantAndrea PoloNo ratings yet
- pcx093 LaudanineDocument13 pagespcx093 LaudanineLulu SetiyabudiNo ratings yet
- 2 Chất Màu Trong Củ DềnDocument12 pages2 Chất Màu Trong Củ Dềnchanchan101103No ratings yet
- Poly (3-Hydroxybutyrate) (PHB) Synthesis by Recombinant Biosynthesis Genes: Effect of Various Carbon and Nitrogen SourcesDocument9 pagesPoly (3-Hydroxybutyrate) (PHB) Synthesis by Recombinant Biosynthesis Genes: Effect of Various Carbon and Nitrogen SourceshafsaNo ratings yet
- La Vie en Rose'': Biosynthesis, Sources, and Applications of Betalain PigmentsDocument16 pagesLa Vie en Rose'': Biosynthesis, Sources, and Applications of Betalain PigmentsNemes BiancaNo ratings yet
- Structural Implications On Color, Xuorescence, and Antiradical Activity in BetalainsDocument12 pagesStructural Implications On Color, Xuorescence, and Antiradical Activity in BetalainsNeyder PantojaNo ratings yet
- Biosynthesis of Medicinal Tropane Alkaloids in Yeast: ArticleDocument26 pagesBiosynthesis of Medicinal Tropane Alkaloids in Yeast: ArticleChandra ShekharNo ratings yet
- LWT - Food Science and Technology: Mohammad Imtiyaj Khan, P.S.C. Sri Harsha, P. Giridhar, G.A. RavishankarDocument9 pagesLWT - Food Science and Technology: Mohammad Imtiyaj Khan, P.S.C. Sri Harsha, P. Giridhar, G.A. RavishankarJoulesNo ratings yet
- Original Article Betalains: Properties, Sources, Applications, and Stability - A ReviewDocument12 pagesOriginal Article Betalains: Properties, Sources, Applications, and Stability - A ReviewTrương Thị Kim LyNo ratings yet
- Betalains - Properties, Sources, Applications, and Stability - A Review PDFDocument12 pagesBetalains - Properties, Sources, Applications, and Stability - A Review PDFBiyaya San PedroNo ratings yet
- Differential Expression ... Unicellular ChlorophyteDocument7 pagesDifferential Expression ... Unicellular ChlorophyteAJITHKUMARNo ratings yet
- Fowler Etal. Melanización de FlavonoidesDocument8 pagesFowler Etal. Melanización de FlavonoidesSaila Viridiana CazaresNo ratings yet
- B-Cyclodextrin Production by The Cyclodextrin Glucanotransferase From Paenibacillus Illinoisensis ZY-08: Cloning, Purification, and PropertiesDocument9 pagesB-Cyclodextrin Production by The Cyclodextrin Glucanotransferase From Paenibacillus Illinoisensis ZY-08: Cloning, Purification, and PropertiesrezqNo ratings yet
- Cytochrome P450 and O-Methyltransferase Catalyze The Final Steps in The Biosynthesis of The Anti-Addictive Alkaloid Ibogaine From Tabernanthe IbogaDocument14 pagesCytochrome P450 and O-Methyltransferase Catalyze The Final Steps in The Biosynthesis of The Anti-Addictive Alkaloid Ibogaine From Tabernanthe IbogaternyNo ratings yet
- Sleat, Mah - Clostridium Populeti Sp. Nov., A Cellulolytic Species From A Woody-Biomass DigestorDocument4 pagesSleat, Mah - Clostridium Populeti Sp. Nov., A Cellulolytic Species From A Woody-Biomass Digestorsukanta60No ratings yet
- Liu IIIDocument8 pagesLiu IIIaji subaktiNo ratings yet
- Xu2015 Article RapidGenerationAndAnalysisOfExDocument13 pagesXu2015 Article RapidGenerationAndAnalysisOfExIrsyadKamalNo ratings yet
- Stability of Betacyanin Pigments FROM RED PURPLE PITAYA FRUIT (Hylocereus Polyrhizus) : Influence of PH, Temperature, Metal Ions and Ascorbic AcidDocument5 pagesStability of Betacyanin Pigments FROM RED PURPLE PITAYA FRUIT (Hylocereus Polyrhizus) : Influence of PH, Temperature, Metal Ions and Ascorbic AcidGufa Bagus PamungkasNo ratings yet
- Molecules: Anthocyanin Pigments: Beyond AestheticsDocument17 pagesMolecules: Anthocyanin Pigments: Beyond AestheticsNguyen Thi Thuy Duong B1606703No ratings yet
- Polymers of 3-Hydroxyacids in Plants: Enea Rezzonico, Laurence Moire & Yves PoirierDocument6 pagesPolymers of 3-Hydroxyacids in Plants: Enea Rezzonico, Laurence Moire & Yves PoirierhafsaNo ratings yet
- J. Biol. Chem.-1999-Garc - A-29228-41Document15 pagesJ. Biol. Chem.-1999-Garc - A-29228-41Héctor AguayoNo ratings yet
- Zhao 2019Document10 pagesZhao 2019Mishel PazmiñoNo ratings yet
- 10 CarotenoidesingmetabolicaDocument7 pages10 CarotenoidesingmetabolicaIris MoralesNo ratings yet
- 1 s2.0 S0176161712003963 MainDocument9 pages1 s2.0 S0176161712003963 MainQuenguan FranklinNo ratings yet
- Multiple B-Ketothiolases Mediate Poly (B-Hydroxyalkanoate) Copolymer Synthesis in Ralstonia EutrophaDocument9 pagesMultiple B-Ketothiolases Mediate Poly (B-Hydroxyalkanoate) Copolymer Synthesis in Ralstonia EutrophaKenneth Jack Zegarra HuamánNo ratings yet
- Characterization of Polyhydroxyalkanoates Accumulated by A Moderately Halophilic Salt Pan Isolate Bacillus Megaterium Strain H16Document10 pagesCharacterization of Polyhydroxyalkanoates Accumulated by A Moderately Halophilic Salt Pan Isolate Bacillus Megaterium Strain H16Luis RodriguezNo ratings yet
- 0 s2.0 S0168165622000268 MainDocument8 pages0 s2.0 S0168165622000268 MainOmar KHELILNo ratings yet
- 39827-Article Text-191359-1-10-20181213Document10 pages39827-Article Text-191359-1-10-20181213Deena SriyazzNo ratings yet
- Colors Health EffectsDocument8 pagesColors Health EffectsemilyNo ratings yet
- Original Article Study On Efficiency of Betacyanin Extraction From Red BeetrootsDocument6 pagesOriginal Article Study On Efficiency of Betacyanin Extraction From Red BeetrootsDehison Guerrero PurisNo ratings yet
- A Metal-Free Blue Chromophore Derived From Plant PigmentsDocument8 pagesA Metal-Free Blue Chromophore Derived From Plant PigmentsLe Ho Chon DuyenNo ratings yet
- 94169-Article Text-241718-1-10-20130912Document13 pages94169-Article Text-241718-1-10-20130912Gaurav ChauhanNo ratings yet
- Art.3 BiosistesisDocument11 pagesArt.3 BiosistesisAngela GuidoNo ratings yet
- Carotene Hydroxylase Activity Determines The Levels of BothDocument11 pagesCarotene Hydroxylase Activity Determines The Levels of BothAna Clara FerreiraNo ratings yet
- Stability of Betalain Pigments From Red BeetrootDocument12 pagesStability of Betalain Pigments From Red BeetrootGovindanayagi PattabiramanNo ratings yet
- Biodegradable Plastic PolymerDocument9 pagesBiodegradable Plastic PolymerAnida Maria Moraes GomesNo ratings yet
- 2 - Genetics and Molecular Biology of Carotenoid Pigment (1996)Document10 pages2 - Genetics and Molecular Biology of Carotenoid Pigment (1996)Gabriel Fernando ParraNo ratings yet
- Variation in Tropane Alkaloid Accumulation Within The Solanaceae and Strategies For Its ExploitationDocument6 pagesVariation in Tropane Alkaloid Accumulation Within The Solanaceae and Strategies For Its ExploitationJuan AspilcuetaNo ratings yet
- Unang Shiono NPR 2019b PDFDocument8 pagesUnang Shiono NPR 2019b PDFazmi azhariNo ratings yet
- Porphyrium Culture Media Optimizationfor GrowthDocument9 pagesPorphyrium Culture Media Optimizationfor GrowthAnonymous Wcj4C3jNo ratings yet
- Higuchi Takeuchi2019Document10 pagesHiguchi Takeuchi2019Adrian Tun PantiNo ratings yet
- Jackman 1996Document66 pagesJackman 199612545343No ratings yet
- An β-1,4-Xylanase with Exo-Enzyme Activity Produced by Paenibacillus xylanilyticus KJ-03 and its cloning and characterizationDocument8 pagesAn β-1,4-Xylanase with Exo-Enzyme Activity Produced by Paenibacillus xylanilyticus KJ-03 and its cloning and characterizationAlonsoNo ratings yet
- Betacyanins in Fruits From Red-Purple Pitaya, Hylocereus Polyrhizus (Weber) Britton & RoseDocument6 pagesBetacyanins in Fruits From Red-Purple Pitaya, Hylocereus Polyrhizus (Weber) Britton & RoseBiyaya San PedroNo ratings yet
- Atropa Belladonna Generated From Hairy Roots PMT and h6h GeneDocument6 pagesAtropa Belladonna Generated From Hairy Roots PMT and h6h GeneHumiopNo ratings yet
- Casaca Da Pitaya ExtraçãoDocument6 pagesCasaca Da Pitaya ExtraçãoPatricia VellanoNo ratings yet
- Food ChemistryDocument5 pagesFood ChemistryKesya HutagaolNo ratings yet
- Pgac 274Document12 pagesPgac 274Gustavo HoppeNo ratings yet
- Chlorophylls and Carotenoids Pigments of Photosynthetic Biomembranes PDFDocument33 pagesChlorophylls and Carotenoids Pigments of Photosynthetic Biomembranes PDFAhmad Hassan ChNo ratings yet
- Vishnuvardhan Reddy 2008 Isolation of PHA Bacteria From SewagesludgeDocument7 pagesVishnuvardhan Reddy 2008 Isolation of PHA Bacteria From SewagesludgeASWINI J SNo ratings yet
- Polyamine Biosynthesis in Arabidopsis ThalianaDocument14 pagesPolyamine Biosynthesis in Arabidopsis ThalianamNo ratings yet
- Production of Xylanase in Transgenic Tobacco For IndustrialDocument12 pagesProduction of Xylanase in Transgenic Tobacco For IndustrialLee BiRonNo ratings yet
- Zanoni Et Al 2020Document8 pagesZanoni Et Al 2020Lorenth ArgüellesNo ratings yet
- Hong 2005Document11 pagesHong 2005tea.zakicNo ratings yet
- Anthocyanin Production by Over-Expression of GrapeDocument9 pagesAnthocyanin Production by Over-Expression of GrapeSudarshanee GeekiyanageNo ratings yet
- New Recombinant Escherichia Coli Strain Tailored For The ProductionDocument6 pagesNew Recombinant Escherichia Coli Strain Tailored For The ProductionAndresMauricio MontesGonzalezNo ratings yet
- Cloning and Sequencing of Gene of and Relationship of Its Product To Some Other Chitinases and Chitinase-Like ProteinsDocument8 pagesCloning and Sequencing of Gene of and Relationship of Its Product To Some Other Chitinases and Chitinase-Like ProteinsFrontiersNo ratings yet
- Lycopene Synthesis and Related Gene Expression in Pummelo Pulp Increased in Shade-Grown FruitDocument7 pagesLycopene Synthesis and Related Gene Expression in Pummelo Pulp Increased in Shade-Grown Fruitnhc_king_proNo ratings yet
- Bio 100 A Virtual Labs Unit One and TwoDocument14 pagesBio 100 A Virtual Labs Unit One and TwoTammy Smith92% (12)
- BIOLOGY 1 MytopschoolsDocument92 pagesBIOLOGY 1 MytopschoolsChinedum danielNo ratings yet
- Lab 2Document11 pagesLab 2Han Hui Chong0% (1)
- Biology Edexcel Unit 2 Model AnswersDocument2 pagesBiology Edexcel Unit 2 Model Answers--------82% (22)
- Final Demo DLP Abenio MaristellaDocument15 pagesFinal Demo DLP Abenio Maristellaapi-668289592No ratings yet
- Borowitzka Et Al 2016 The Physiology of Microalgae, The Cell Cycle of MicroalgaeDocument44 pagesBorowitzka Et Al 2016 The Physiology of Microalgae, The Cell Cycle of MicroalgaeKarinaNo ratings yet
- Test Bank For Genetics and Genomics in Nursing and Health Care 2nd Edition by BeeryDocument9 pagesTest Bank For Genetics and Genomics in Nursing and Health Care 2nd Edition by BeeryFranklin Roberts100% (39)
- Full Download Human Heredity Principles and Issues 10th Edition Michael Cummings Solutions ManualDocument36 pagesFull Download Human Heredity Principles and Issues 10th Edition Michael Cummings Solutions Manualwalmmidgetoin38o100% (29)
- 2ND Year CytogeneticsDocument11 pages2ND Year Cytogenetics뷸한연No ratings yet
- Judul Tugas Studi MandiriDocument18 pagesJudul Tugas Studi MandiriHasan Assya IdahNo ratings yet
- Analysis of Zinc in Human Blood Using Flame Atomic Absorption Spectroscopy (FAAS)Document3 pagesAnalysis of Zinc in Human Blood Using Flame Atomic Absorption Spectroscopy (FAAS)sujonosuputroNo ratings yet
- Malignant Transformation of Hymenolepis Nana in A Human HostDocument9 pagesMalignant Transformation of Hymenolepis Nana in A Human HostvictoriaNo ratings yet
- Science 8 - Q4Document15 pagesScience 8 - Q4Lance Delos SantosNo ratings yet
- Module 3 - Gen Bio 1 - MidtermDocument7 pagesModule 3 - Gen Bio 1 - MidtermAngel Cuacko GacmatanNo ratings yet
- Introduction To Cell and Tissue CultureDocument314 pagesIntroduction To Cell and Tissue CulturereddygonaNo ratings yet
- Chapter 5: The Mitotic Cell CycleDocument7 pagesChapter 5: The Mitotic Cell CycleWei Jern0% (1)
- Cell Kinetics Chapter 6Document56 pagesCell Kinetics Chapter 6Sanjay Kumar73% (11)
- Classification of OncogenesDocument20 pagesClassification of Oncogenes18B236 - NIVYA SUBULAXMI MNo ratings yet
- The Cell Cycle and Cell ProliferationDocument49 pagesThe Cell Cycle and Cell ProliferationLeah BeningtonNo ratings yet
- Cell Stem - November 2012Document148 pagesCell Stem - November 2012candle stickNo ratings yet
- Human Heredity Principles and Issues 11th Edition Cummings Solution ManualDocument56 pagesHuman Heredity Principles and Issues 11th Edition Cummings Solution Manualmichael100% (20)
- Module 2 Cell Cycle and Cell DivisionDocument9 pagesModule 2 Cell Cycle and Cell DivisionBalolot RalphNo ratings yet
- IB BIOLOGY TOPIC 1 QuestionsDocument3 pagesIB BIOLOGY TOPIC 1 QuestionsSarah Mason100% (1)
- Biology 2019Document32 pagesBiology 2019Rinchen DemaNo ratings yet
- Lampl - 1992 - Saltation and StasisDocument4 pagesLampl - 1992 - Saltation and StasistechnopsychNo ratings yet
- Extracted Pages From Cambridge International AS and A Level Biology Students Book 2nd Edition (C. J. Clegg, Geoff Goodwin) (Z-Lib - Org) 1Document5 pagesExtracted Pages From Cambridge International AS and A Level Biology Students Book 2nd Edition (C. J. Clegg, Geoff Goodwin) (Z-Lib - Org) 1BonnyNo ratings yet
- As BiologyDocument127 pagesAs BiologyRob Gomez100% (5)
- Cell DivisionDocument13 pagesCell DivisionNaimah HadiqahNo ratings yet
- Cell CycleDocument18 pagesCell CyclewaveafterwaveNo ratings yet
- Biology Class 10 Icse Boards!Document10 pagesBiology Class 10 Icse Boards!nark.jf4No ratings yet
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (6)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (28)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeRating: 2 out of 5 stars2/5 (1)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (1)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (404)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (81)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningFrom EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningRating: 4 out of 5 stars4/5 (3)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)From EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)No ratings yet
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Marshmallow Test: Mastering Self-ControlFrom EverandThe Marshmallow Test: Mastering Self-ControlRating: 4.5 out of 5 stars4.5/5 (58)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Dark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingFrom EverandDark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingRating: 4 out of 5 stars4/5 (1138)
- Troubled: A Memoir of Foster Care, Family, and Social ClassFrom EverandTroubled: A Memoir of Foster Care, Family, and Social ClassRating: 4.5 out of 5 stars4.5/5 (26)