You might also like
- 2017, Elife, RiceDocument29 pages2017, Elife, RiceOLGA MARÍNNo ratings yet
- Transcriptional Regulation of The ER Stress-Inducible Gene Sec16B in Neuro2a CellsDocument10 pagesTranscriptional Regulation of The ER Stress-Inducible Gene Sec16B in Neuro2a CellsJulia MouraNo ratings yet
- Differential Expression of LNC RNADocument12 pagesDifferential Expression of LNC RNAnikhita.chirraNo ratings yet
- Ancient Endo-siRNA Pathways Reveal New Tricks in Gene RegulationDocument13 pagesAncient Endo-siRNA Pathways Reveal New Tricks in Gene RegulationAntarToumaNo ratings yet
- Nature 11247Document18 pagesNature 11247Cristina PavelNo ratings yet
- Jurnal GonadDocument7 pagesJurnal GonadMr lonelyNo ratings yet
- Ref 3Document13 pagesRef 3Rohit MaliNo ratings yet
- Dicing Bodies1Document6 pagesDicing Bodies1Zain BaderNo ratings yet
- 39 564 1 PBDocument13 pages39 564 1 PBpentingskripsiNo ratings yet
- RNA InterferenceDocument26 pagesRNA InterferenceAjayChandrakarNo ratings yet
- The RNA-Induced SilencingDocument5 pagesThe RNA-Induced Silencingchavi1310No ratings yet
- Sirna and Mirna Processing: New Functions For Cajal Bodies: Olga Pontes and Craig S PikaardDocument7 pagesSirna and Mirna Processing: New Functions For Cajal Bodies: Olga Pontes and Craig S PikaardChristian Moises CasasNo ratings yet
- RNA Interference in J774 Macrophages Reveals A Role For Coronin 1 in Mycobacterial Trafficking But Not in Actin-Dependent ProcessesDocument11 pagesRNA Interference in J774 Macrophages Reveals A Role For Coronin 1 in Mycobacterial Trafficking But Not in Actin-Dependent ProcessesAbdulaziz AlharbiNo ratings yet
- What Were The Most Important Discoveries in Our Reviewed PaperDocument10 pagesWhat Were The Most Important Discoveries in Our Reviewed PaperchrisNo ratings yet
- 2015 Comparative Analysis of The Small RNA Transcriptomes of Pinus Contorta and Oryza SativaDocument15 pages2015 Comparative Analysis of The Small RNA Transcriptomes of Pinus Contorta and Oryza Sativaplanhigion06No ratings yet
- RNA Interference Class NotesDocument4 pagesRNA Interference Class NotesmanaleepandyaNo ratings yet
- Genome Res.-2012-Stadler-2418-26Document10 pagesGenome Res.-2012-Stadler-2418-26frostyNo ratings yet
- tRNA-derived Small RNAs Target Transposable Element TranscriptsDocument11 pagestRNA-derived Small RNAs Target Transposable Element TranscriptsMan Ba SotzNo ratings yet
- Gene Silencing by Small RNAs in Invertebrates and VertebratesDocument7 pagesGene Silencing by Small RNAs in Invertebrates and VertebratesLateecka R KulkarniNo ratings yet
- Isolation and Expression of Two Distinct Sox8 Genes in Mudloach (Misgurnus Anguillicaudatus)Document16 pagesIsolation and Expression of Two Distinct Sox8 Genes in Mudloach (Misgurnus Anguillicaudatus)josekinNo ratings yet
- Genome Res.-2021-Hori-1971-82Document13 pagesGenome Res.-2021-Hori-1971-82TiffanyNo ratings yet
- 4 A To I Editing 2003Document4 pages4 A To I Editing 2003milenerato2240No ratings yet
- Fmolb 08 708984Document5 pagesFmolb 08 708984Aaser Ammar Ali HassanNo ratings yet
- Diencephalic and Neuropeptidergic Dysfunction in Zebrafish With Autism Risk MutationsDocument26 pagesDiencephalic and Neuropeptidergic Dysfunction in Zebrafish With Autism Risk MutationszbtyvsknfdNo ratings yet
- Sharma-Vakhlu2021 Article EvolutionAndBiologyOfCRISPRSysDocument22 pagesSharma-Vakhlu2021 Article EvolutionAndBiologyOfCRISPRSysMARÍA ALEJANDRA FLORIAN CORTESNo ratings yet
- Plant Virus Interaction - OrelvisDocument66 pagesPlant Virus Interaction - OrelvisJoseph Rafael Aguayo RuizNo ratings yet
- Oligopeptide Polyarginine Mediated Delivery of Short Interfering RNA To Cells For Gene SilencingDocument7 pagesOligopeptide Polyarginine Mediated Delivery of Short Interfering RNA To Cells For Gene SilencingPaul HartingNo ratings yet
- Genetic ResearchDocument14 pagesGenetic ResearchDr LipseyNo ratings yet
- CDE 1 Affects Chromosome Segregation Through Uridy PDFDocument14 pagesCDE 1 Affects Chromosome Segregation Through Uridy PDFcgilgasNo ratings yet
- 1743 422X 8 178Document14 pages1743 422X 8 178Kalyan Kumar PasumarthyNo ratings yet
- Elbashir Et Al. 2001Document12 pagesElbashir Et Al. 2001Benedikt EngelNo ratings yet
- Testosterone - Action3Document18 pagesTestosterone - Action3Koul RodoNo ratings yet
- Trans-kingdomCross-Talk. SmallRNAsontheMove - 2014Document7 pagesTrans-kingdomCross-Talk. SmallRNAsontheMove - 2014edNo ratings yet
- GKW 139Document13 pagesGKW 139azmul nurNo ratings yet
- Malik Et Al - 2021Document21 pagesMalik Et Al - 2021indibiochemNo ratings yet
- Biochemical Activity of Is RDR6Document8 pagesBiochemical Activity of Is RDR6Quanan HuNo ratings yet
- Nucl. Acids Res.-2011-Sakurai-1510-25Document16 pagesNucl. Acids Res.-2011-Sakurai-1510-25farhana rahmanNo ratings yet
- Gene Regulation of Mammalian Long Non Coding RNA: Heeyoun BunchDocument15 pagesGene Regulation of Mammalian Long Non Coding RNA: Heeyoun Bunchmehdi_mohebi2009No ratings yet
- Cross-Mapping MicroRNA IdentificationDocument8 pagesCross-Mapping MicroRNA IdentificationRicardo GoreNo ratings yet
- CRISPR HandbookDocument18 pagesCRISPR Handbookgoucloud100% (1)
- A Antisense Technology 1Document7 pagesA Antisense Technology 1Gourav JainNo ratings yet
- Functional Genomics Is A Field ofDocument6 pagesFunctional Genomics Is A Field ofGuru RaviNo ratings yet
- RNA Silencing in Chlamydomonas PDFDocument16 pagesRNA Silencing in Chlamydomonas PDFGerman GodiNo ratings yet
- RNAi Discovery and Mechanism in 40 CharactersDocument6 pagesRNAi Discovery and Mechanism in 40 Charactersaman jaiswalNo ratings yet
- 2016-Reevaluation of The Roles of DROSHA, Exportin 5, and DICER in MiRNA BiogenesisDocument9 pages2016-Reevaluation of The Roles of DROSHA, Exportin 5, and DICER in MiRNA BiogenesisAntarToumaNo ratings yet
- Non-Coding RNA Research: He Chen, Ge Shan TDocument7 pagesNon-Coding RNA Research: He Chen, Ge Shan TAsim MughalNo ratings yet
- Castañeda 2016Document16 pagesCastañeda 2016papahojaloveNo ratings yet
- J Biol Chem 2003 Seshadri VDocument8 pagesJ Biol Chem 2003 Seshadri VHeiri StarkNo ratings yet
- Genome-Wide Analyses Across Viridiplantae Reveal TDocument12 pagesGenome-Wide Analyses Across Viridiplantae Reveal TVictor CostaNo ratings yet
- Resistance Response of Indonesian Native Orchids To Odontoglossum Ringspot VirusDocument7 pagesResistance Response of Indonesian Native Orchids To Odontoglossum Ringspot VirusRafiq FananiNo ratings yet
- This Week in Nature and Science 1Document2 pagesThis Week in Nature and Science 1Irene SaezNo ratings yet
- Ponting, Et Al 2009. Evolution and Functions of Long Noncoding RNAs.Document13 pagesPonting, Et Al 2009. Evolution and Functions of Long Noncoding RNAs.J Alberto LucasNo ratings yet
- Clemens, J.C. (2000) PDFDocument6 pagesClemens, J.C. (2000) PDFLateecka R KulkarniNo ratings yet
- Fmrp-Mediated Axonal Delivery of Mir-181D Regulates Axon Elongation by Locally Targeting Map1B and Calm1Document15 pagesFmrp-Mediated Axonal Delivery of Mir-181D Regulates Axon Elongation by Locally Targeting Map1B and Calm1dupuytrenNo ratings yet
- Gene PredictionDocument15 pagesGene PredictionRaghav SureshNo ratings yet
- Genome Res.-2007-He-1471-7Document8 pagesGenome Res.-2007-He-1471-7Richard Icaro EsparzaNo ratings yet
- 8 Sigmas2003Document7 pages8 Sigmas2003AsmaNo ratings yet
- 47 FullDocument7 pages47 Fulldrscribd25No ratings yet
- 2010 Chiang MamalianmiRNASDocument19 pages2010 Chiang MamalianmiRNASDejanete RodriguesNo ratings yet
- Terms and ConditionsDocument2 pagesTerms and ConditionsLateecka R KulkarniNo ratings yet
- Terms and ConditionsDocument1 pageTerms and ConditionsSandeepNo ratings yet
- International Archives of Medicine: Health Consequences of Cricket - View From South AsiaDocument16 pagesInternational Archives of Medicine: Health Consequences of Cricket - View From South AsiaLateecka R KulkarniNo ratings yet
- BMC Public HealthDocument17 pagesBMC Public HealthLateecka R KulkarniNo ratings yet
- 5 Minute Workout - Brazillian Butt LiftDocument3 pages5 Minute Workout - Brazillian Butt LiftLateecka R KulkarniNo ratings yet
- 1471 2458 13 683Document19 pages1471 2458 13 683Lateecka R KulkarniNo ratings yet
- 1756 0500 6 296Document13 pages1756 0500 6 296Lateecka R KulkarniNo ratings yet
- Inner Ring Groove & Land Grinding ProcessDocument38 pagesInner Ring Groove & Land Grinding ProcessLateecka R Kulkarni100% (1)
- Guides - Non-Clzroom Use (055) : 90 Note 187pDocument187 pagesGuides - Non-Clzroom Use (055) : 90 Note 187pLateecka R KulkarniNo ratings yet
- 1756 0500 6 294Document20 pages1756 0500 6 294Lateecka R KulkarniNo ratings yet
- 1475 2875 12 262Document33 pages1475 2875 12 262Lateecka R KulkarniNo ratings yet
- 1754 6834 6 108Document27 pages1754 6834 6 108Lateecka R KulkarniNo ratings yet
- 1471 2334 13 347Document9 pages1471 2334 13 347Lateecka R KulkarniNo ratings yet
- 1471 2458 13 686Document18 pages1471 2458 13 686Lateecka R KulkarniNo ratings yet
- BMC GenomicsDocument25 pagesBMC GenomicsLateecka R KulkarniNo ratings yet
- 1471 2164 14 504Document31 pages1471 2164 14 504Lateecka R KulkarniNo ratings yet
- 1754 6834 6 108Document27 pages1754 6834 6 108Lateecka R KulkarniNo ratings yet
- 1471 2334 13 347Document9 pages1471 2334 13 347Lateecka R KulkarniNo ratings yet
- World Journal of Emergency SurgeryDocument9 pagesWorld Journal of Emergency SurgeryLateecka R KulkarniNo ratings yet
- 1532 429X 15 65Document26 pages1532 429X 15 65Lateecka R KulkarniNo ratings yet
- 1471 2377 13 98Document10 pages1471 2377 13 98Lateecka R KulkarniNo ratings yet
- Lipids in Health and DiseaseDocument16 pagesLipids in Health and DiseaseLateecka R KulkarniNo ratings yet
- 1477 7819 11 169Document14 pages1477 7819 11 169Lateecka R KulkarniNo ratings yet
- 1471 2377 13 97Document15 pages1471 2377 13 97Lateecka R KulkarniNo ratings yet
- 1743 422X 10 245Document27 pages1743 422X 10 245Lateecka R KulkarniNo ratings yet
- 1471 244X 13 197Document38 pages1471 244X 13 197Lateecka R KulkarniNo ratings yet
- 1471 2148 13 158Document29 pages1471 2148 13 158Lateecka R KulkarniNo ratings yet
- 1479 5876 11 180Document22 pages1479 5876 11 180Lateecka R KulkarniNo ratings yet
- BMC BioinformaticsDocument24 pagesBMC BioinformaticsLateecka R KulkarniNo ratings yet
- 1471 244X 13 198Document28 pages1471 244X 13 198Lateecka R KulkarniNo ratings yet
- BS 08545-2013Document97 pagesBS 08545-2013Stanislav PerevezentsevNo ratings yet
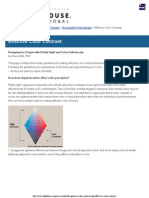
- Lighthouse International - Effective Color ContrastDocument5 pagesLighthouse International - Effective Color ContrastVaishnavi JayakumarNo ratings yet
- Evolutiontestreviewdoc 2015Document5 pagesEvolutiontestreviewdoc 2015api-265621022No ratings yet
- Soil Classification (Taxonomy)Document58 pagesSoil Classification (Taxonomy)Quinnee VallejosNo ratings yet
- Competitive Intelligence AcceraDocument29 pagesCompetitive Intelligence AcceranarenebiowebNo ratings yet
- Kompetisi Sains Medalyst Bahasa Inggris SMPDocument19 pagesKompetisi Sains Medalyst Bahasa Inggris SMPNindia Ayu FaridshaNo ratings yet
- Geriatric PanchakarmaDocument24 pagesGeriatric Panchakarmaksr prasadNo ratings yet
- b2601 Course Outline and Practical Manual - 2020-2021Document24 pagesb2601 Course Outline and Practical Manual - 2020-2021sispulieNo ratings yet
- Notes - Growth and DevelopmentDocument4 pagesNotes - Growth and DevelopmentNathan mwapeNo ratings yet
- ContinueDocument2 pagesContinueShallu ManchandaNo ratings yet
- 10.3 Gene Pools and SpeciationstudentnotesDocument4 pages10.3 Gene Pools and SpeciationstudentnotesMoahmed Mahmoud IB15A 363KAGYNo ratings yet
- Bio-Diversity Uses Threats and ConservationDocument32 pagesBio-Diversity Uses Threats and ConservationRoshin VargheseNo ratings yet
- Allplex SARS-CoV-2 Variant I AssaysDocument1 pageAllplex SARS-CoV-2 Variant I AssaysadibyanNo ratings yet
- Myofascial Meridians PDFDocument9 pagesMyofascial Meridians PDFPedja Ley100% (3)
- P3. Membrane TransportDocument6 pagesP3. Membrane TransportCrisNo ratings yet
- Application of Edible Films and Coatings On MeatsDocument2 pagesApplication of Edible Films and Coatings On MeatsAnasZeidNo ratings yet
- Individual Differences and Emotional IntelligenceDocument91 pagesIndividual Differences and Emotional IntelligenceSREEKUTTY K MNo ratings yet
- Chapter 5 - DNADocument8 pagesChapter 5 - DNAzsuzanna100% (1)
- Microfossils Description Paper: Palaeontology Practical: KalpanadekakalitaDocument32 pagesMicrofossils Description Paper: Palaeontology Practical: Kalpanadekakalitarikalave ramanNo ratings yet
- Lichens-Introduction, Classification, Structure, Reproduction and Economic Importance by Suresh Prashad Bhatta (M. Sc. Botany)Document11 pagesLichens-Introduction, Classification, Structure, Reproduction and Economic Importance by Suresh Prashad Bhatta (M. Sc. Botany)Suresh BhattaNo ratings yet
- Soal Pat Inggris Kelas 7 - 2021Document9 pagesSoal Pat Inggris Kelas 7 - 2021Bella Septiani FaryanNo ratings yet
- Presentation On Hydration PDFDocument17 pagesPresentation On Hydration PDFManuel CastroNo ratings yet
- Karakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiDocument7 pagesKarakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiAlexanderNo ratings yet
- Chapter 1 of Blue MindDocument23 pagesChapter 1 of Blue MindSouthern California Public Radio100% (1)
- Rankem SolventsDocument6 pagesRankem SolventsAmmar MohsinNo ratings yet
- Erl mf2 PDFDocument2 pagesErl mf2 PDFJorgeNo ratings yet
- AP Bio Lab 1Document5 pagesAP Bio Lab 1Kiran ShilaNo ratings yet
- Biological Classification Class 11 Notes BiologyDocument6 pagesBiological Classification Class 11 Notes BiologyVyjayanthiNo ratings yet
- Dr. Morse's Herbal Formulation ListDocument10 pagesDr. Morse's Herbal Formulation ListTom50% (2)
- Morphological Characterization of Pichia Guilliermondii and Saccharomyces Cerevisiae Yeast and Their Effects On Adherence of IntesDocument7 pagesMorphological Characterization of Pichia Guilliermondii and Saccharomyces Cerevisiae Yeast and Their Effects On Adherence of IntesAnonymous Y83Rx25No ratings yet