You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- 2019 08 21 - Program) OptilasDocument90 pages2019 08 21 - Program) OptilasIvan MontesNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Design and Calibration For A Full-Stokes Imaging PolarimeterDocument11 pagesDesign and Calibration For A Full-Stokes Imaging PolarimeterIvan MontesNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- WarpDrive MathDocument10 pagesWarpDrive Mathaskj78No ratings yet
- Jack Evett Cheng Liu-2500 Solved Problems in Fluid Mechanics and Hydraulics-Mcgraw-Hill 1989Document15 pagesJack Evett Cheng Liu-2500 Solved Problems in Fluid Mechanics and Hydraulics-Mcgraw-Hill 1989Ivan MontesNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Ebook Dho Health Science Updated 8Th Edition Simmers Test Bank Full Chapter PDFDocument28 pagesEbook Dho Health Science Updated 8Th Edition Simmers Test Bank Full Chapter PDFjuliemcintyretzydiengcr100% (10)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Guide to Cataract Surgery RecoveryDocument16 pagesGuide to Cataract Surgery Recoverydokumen kuNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Making of Portrait of Graham and Amanda by Ian Spriggs - CG TUTORIALDocument1 pageMaking of Portrait of Graham and Amanda by Ian Spriggs - CG TUTORIALKennedyKisusiNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Anatomy of Vitreous and RetinaDocument43 pagesAnatomy of Vitreous and RetinatariqNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- 50 Remarkable Facts About ButterfliesDocument59 pages50 Remarkable Facts About Butterfliesdesign designNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Human Facotr Mind MapDocument1 pageHuman Facotr Mind Map张瑜征No ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Lesson Plan 1Document5 pagesLesson Plan 1api-248521749No ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Diagnostic Value of Nystagmus: Spontaneous and Induced Ocular OscillationsDocument5 pagesDiagnostic Value of Nystagmus: Spontaneous and Induced Ocular OscillationsPrincella MonicaNo ratings yet
- Anatomy of Eye AppendigesDocument19 pagesAnatomy of Eye AppendigesSiyaNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Best Colored Contact Lenses To Suit Your Eyes and StyleDocument2 pagesThe Best Colored Contact Lenses To Suit Your Eyes and StyleDarren Noel ChandraNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Cataract Surgery Following Cornea Transplant - American Academy of OphthalmologyDocument2 pagesCataract Surgery Following Cornea Transplant - American Academy of OphthalmologyAlejandro N. Fonseca GrandaNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- 2.4.2.a VisualPerceptionDocument5 pages2.4.2.a VisualPerceptionAnniston NeedhamNo ratings yet
- Halitosis: Common Bad Breath Causes and SolutionsDocument5 pagesHalitosis: Common Bad Breath Causes and Solutionspot poootNo ratings yet
- Cmeseries 28Document88 pagesCmeseries 28ashishNo ratings yet
- Human Sense Organs Lesson PlanDocument152 pagesHuman Sense Organs Lesson PlanJen sag89% (37)
- Eye Sign ExamplesDocument2 pagesEye Sign ExamplesDraganNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- History of PostersDocument27 pagesHistory of PostersSamarth BhagwatNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- FACIAL PROPORTIONS GUIDEDocument19 pagesFACIAL PROPORTIONS GUIDELadunca MihaiNo ratings yet
- Daftar PustakaDocument3 pagesDaftar PustakaDokter LinggauNo ratings yet
- Lens Luxation in DogsDocument4 pagesLens Luxation in DogsBernadhete Gaudia SabiroseNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- June 2020 DNB Question BankDocument3 pagesJune 2020 DNB Question Bankvittal iNo ratings yet
- 21 Steps Revised OptDocument52 pages21 Steps Revised OptRITA OGECHI EZENWOKONo ratings yet
- PHYSICS Class 10th NotesDocument36 pagesPHYSICS Class 10th NotesAwan MalikNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
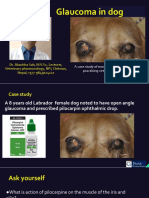
- Glaucoma in Dog A Case Report-Dr. Jibachha SahDocument8 pagesGlaucoma in Dog A Case Report-Dr. Jibachha SahDr.Jibachha Sah,Vet.physician,author, & motivatorNo ratings yet
- Neuro-Ophthalmology: Bethlehem Girma (M.D) Assistant Professor of OphthalmologyDocument117 pagesNeuro-Ophthalmology: Bethlehem Girma (M.D) Assistant Professor of Ophthalmologyhenok birukNo ratings yet
- Jayanth Virtual Retinal DisplayDocument44 pagesJayanth Virtual Retinal Displayapi-3827000100% (5)
- Refraction of LightDocument17 pagesRefraction of LightShari OliverNo ratings yet
- Grading Scales Web PDFDocument2 pagesGrading Scales Web PDFDanielle SangalangNo ratings yet
- Eye Emergency ManualDocument56 pagesEye Emergency Manualgraci91031462296100% (1)
- Course Outline OPSC750 - 201910 PDFDocument4 pagesCourse Outline OPSC750 - 201910 PDFKeshav KafleNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)