You might also like
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Patogenesis Infeksi Virus White Spot (WSV) Pada Udang Windu (Penaeus Monodon Fab.)Document8 pagesPatogenesis Infeksi Virus White Spot (WSV) Pada Udang Windu (Penaeus Monodon Fab.)Tuti Puji LestariNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- B 050113Document5 pagesB 050113Biodiversitas, etc100% (5)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Cloning and Tissue-Specific Expression of Cytochrome P-450 Aromatase Cdna in Japanese Eel (Anguilla Japonica)Document4 pagesCloning and Tissue-Specific Expression of Cytochrome P-450 Aromatase Cdna in Japanese Eel (Anguilla Japonica)Tuti Puji LestariNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Patogenesis Infeksi Virus White Spot (WSV) Pada Udang Windu (Penaeus Monodon Fab.)Document8 pagesPatogenesis Infeksi Virus White Spot (WSV) Pada Udang Windu (Penaeus Monodon Fab.)Tuti Puji LestariNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- 57-Rothbard Et Al 1990 Clearnace of 17alphaETDocument12 pages57-Rothbard Et Al 1990 Clearnace of 17alphaETTuti Puji LestariNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Atrazine Reduces Reproduction in Japanese Medaka (Oryzias Latipes)Document10 pagesAtrazine Reduces Reproduction in Japanese Medaka (Oryzias Latipes)Tuti Puji LestariNo ratings yet
- Fenotipe Keturunan Pertama Ikan Koi Hasil Ginogenesis Phenotype of The First Gynogenesis Generation of KoiDocument4 pagesFenotipe Keturunan Pertama Ikan Koi Hasil Ginogenesis Phenotype of The First Gynogenesis Generation of KoiTuti Puji LestariNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Steroid Hormone in Serum of Male Mugil CephalusDocument22 pagesSteroid Hormone in Serum of Male Mugil CephalusTuti Puji LestariNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Pengaruh Lama Waktu Pery - Ndaman Induk Di Dalam Larutan Hormon 17 - Metiltestosteron Terhadap Nisbah Kelamin Anak Ikan GapiDocument5 pagesPengaruh Lama Waktu Pery - Ndaman Induk Di Dalam Larutan Hormon 17 - Metiltestosteron Terhadap Nisbah Kelamin Anak Ikan GapiTuti Puji LestariNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Fenotipe Keturunan Pertama Ikan Koi Hasil Ginogenesis Phenotype of The First Gynogenesis Generation of KoiDocument4 pagesFenotipe Keturunan Pertama Ikan Koi Hasil Ginogenesis Phenotype of The First Gynogenesis Generation of KoiTuti Puji LestariNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Cloning and Tissue-Specific Expression of Cytochrome P-450 Aromatase Cdna in Japanese Eel (Anguilla Japonica)Document4 pagesCloning and Tissue-Specific Expression of Cytochrome P-450 Aromatase Cdna in Japanese Eel (Anguilla Japonica)Tuti Puji LestariNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Binder 1Document30 pagesBinder 1Tuti Puji LestariNo ratings yet
- Steroid Hormone in Serum of Male Mugil CephalusDocument22 pagesSteroid Hormone in Serum of Male Mugil CephalusTuti Puji LestariNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Jurnal Khusus Perikanan Dan Mengenai Famili CyprinidDocument16 pagesJurnal Khusus Perikanan Dan Mengenai Famili CyprinidTuti Puji LestariNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- V 23 N 4 A 07Document8 pagesV 23 N 4 A 07Tuti Puji LestariNo ratings yet
- BBM 3A978 3 642 16077 6 2F1Document21 pagesBBM 3A978 3 642 16077 6 2F1Tuti Puji LestariNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Amer. Zool.-1997-KIM-470-81Document12 pagesAmer. Zool.-1997-KIM-470-81Tuti Puji LestariNo ratings yet
- California Abalone AquacultureDocument5 pagesCalifornia Abalone AquacultureTuti Puji LestariNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Oseana - Xxxi (1) 1-7 01Document7 pagesOseana - Xxxi (1) 1-7 01Tuti Puji LestariNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- A 58 Year Old Client Is Admitted With A Diagnosis of Lung CancerDocument9 pagesA 58 Year Old Client Is Admitted With A Diagnosis of Lung CancerNur SanaaniNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Preparing A Family For Childbirth and ParentingDocument5 pagesPreparing A Family For Childbirth and ParentingBern NerquitNo ratings yet
- BDSM Checklist: General InformationDocument6 pagesBDSM Checklist: General Informationmiri100% (1)
- STEVENS DRRR Module 6 Opon Christian Joy ADocument7 pagesSTEVENS DRRR Module 6 Opon Christian Joy AAnthony Bryan Cartujano100% (1)
- Catalogo Baby MarketDocument53 pagesCatalogo Baby Marketmiguel quispeNo ratings yet
- The Good-Enough Sex Model For Couple Sexual SatisfactionDocument13 pagesThe Good-Enough Sex Model For Couple Sexual SatisfactionwernikNo ratings yet
- The Miracle of ChocolateDocument10 pagesThe Miracle of ChocolateAmanda YasminNo ratings yet
- Fluoride Menace in OrissaDocument83 pagesFluoride Menace in OrissaProfessor PrabhatNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Bakery Business PlanDocument31 pagesBakery Business PlanRohit Gupta93% (14)
- Significance of HACCP and SSOP in Food Processing EstablishmentsDocument7 pagesSignificance of HACCP and SSOP in Food Processing EstablishmentselfiraNo ratings yet
- AssignmentDocument2 pagesAssignmentReserva, ArchelleNo ratings yet
- Laboratory Result Report: Requested Test Result Units Reference Value Method ImmunologyDocument1 pageLaboratory Result Report: Requested Test Result Units Reference Value Method ImmunologyYaya ZakariaNo ratings yet
- EOHSP 09 Operational Control ProcedureDocument3 pagesEOHSP 09 Operational Control ProcedureAli ImamNo ratings yet
- Gastritis - Etiology and Diagnosis - UpToDateDocument10 pagesGastritis - Etiology and Diagnosis - UpToDateLizbeth Navarrete SierraNo ratings yet
- 7 DR BookletDocument13 pages7 DR BookletDebraNo ratings yet
- Activity Design (Summer League 2022)Document6 pagesActivity Design (Summer League 2022)Yubert ViosNo ratings yet
- Literature ReviewDocument11 pagesLiterature Reviewapi-582875150No ratings yet
- GP Panel ClinicsDocument9 pagesGP Panel ClinicsKoh WYhowNo ratings yet
- DMF 40 199 PDFDocument14 pagesDMF 40 199 PDFsiraj_khan_13No ratings yet
- Brazilian Sanitary Guide For Cruise ShipsDocument82 pagesBrazilian Sanitary Guide For Cruise ShipsStanislav KozhuharovNo ratings yet
- Ultrafiltration and Its Application in Food Processing: October 2015Document15 pagesUltrafiltration and Its Application in Food Processing: October 2015Doina PolisciucNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Module 3A: Designing Instruction in The Different Learning Delivery ModalitiesDocument30 pagesModule 3A: Designing Instruction in The Different Learning Delivery Modalitiesnel baradi67% (9)
- StoryDocument6 pagesStoryGerard NicholasNo ratings yet
- Transferring A Dependent Patient From Bed To ChairDocument5 pagesTransferring A Dependent Patient From Bed To Chairapi-26570979No ratings yet
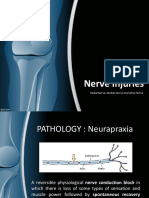
- Nerve Injuries: Radial Nerve, Median Nerve and Ulnar NerveDocument26 pagesNerve Injuries: Radial Nerve, Median Nerve and Ulnar NerveaisiyaahNo ratings yet
- (Cô Nguyêt CA) .Jump Ielts - Writing Task 1 & 2 (Abazar Alizadeh)Document140 pages(Cô Nguyêt CA) .Jump Ielts - Writing Task 1 & 2 (Abazar Alizadeh)Hong HoaNo ratings yet
- Tonsillectomy and Adenoidectomy - Pediatric Clinics of North AmericaDocument13 pagesTonsillectomy and Adenoidectomy - Pediatric Clinics of North AmericayosefNo ratings yet
- Abc Ven 2020Document81 pagesAbc Ven 2020CorneLia JacintaNo ratings yet
- Nutrition Case Study Presentation Slides - Jacob NewmanDocument27 pagesNutrition Case Study Presentation Slides - Jacob Newmanapi-283248618No ratings yet
- Phytophthora InfestansDocument10 pagesPhytophthora Infestansvas2000No ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)