You might also like
- Marine Conservation Biology: The Science of Maintaining the Sea's BiodiversityFrom EverandMarine Conservation Biology: The Science of Maintaining the Sea's BiodiversityRating: 5 out of 5 stars5/5 (1)
- The State of The Science - Forage Fish in The California Current.Document20 pagesThe State of The Science - Forage Fish in The California Current.PewEnvironmentGroupNo ratings yet
- Stephen Gonzales Revised Final 1Document6 pagesStephen Gonzales Revised Final 1api-582795028No ratings yet
- Global Threat to Coral Reefs Harms Poorest NationsDocument3 pagesGlobal Threat to Coral Reefs Harms Poorest NationsSamar AhmadNo ratings yet
- Jhered Esq088 FullDocument14 pagesJhered Esq088 FullCarlos Pizarro GomezNo ratings yet
- Mascareñas-Osorio Et Al. - 2011 - Checklist of Conspicuous Reef Fishes of the Bahía .PDFDocument9 pagesMascareñas-Osorio Et Al. - 2011 - Checklist of Conspicuous Reef Fishes of the Bahía .PDFDaniel OviedoNo ratings yet
- researchDocument7 pagesresearchChristine CuencaNo ratings yet
- Asociaciones PecesDocument13 pagesAsociaciones PecesjorgelopezrochaNo ratings yet
- BioScience 2005 Allan 1041 51Document11 pagesBioScience 2005 Allan 1041 51amandamendaNo ratings yet
- Contador Et Al. 2012Document13 pagesContador Et Al. 2012Cata VerdugoNo ratings yet
- UntitledDocument8 pagesUntitledKyle Keen TaerNo ratings yet
- Habitat Suitability and Anthropogenic Correlates of Neotropical River Otter (LontraDocument11 pagesHabitat Suitability and Anthropogenic Correlates of Neotropical River Otter (LontraMariaVictoriaCastañedaUsugaNo ratings yet
- Testing The Generality of The Trophic Cascade Paradigm For Sea Otters: A Case Study With Kelp Forests in Northern Washington, USADocument18 pagesTesting The Generality of The Trophic Cascade Paradigm For Sea Otters: A Case Study With Kelp Forests in Northern Washington, USAapi-648008046No ratings yet
- Sustainability of Capture of Fish Bycatch in The Prawn Trawling in The Northeastern BrazilDocument10 pagesSustainability of Capture of Fish Bycatch in The Prawn Trawling in The Northeastern BrazilCarlos JúniorNo ratings yet
- Literature Review Sustainable DevelopmentDocument5 pagesLiterature Review Sustainable DevelopmentJade NelsonNo ratings yet
- Insights into genetic diversity and group composition of Atlantic white-sided dolphinsDocument9 pagesInsights into genetic diversity and group composition of Atlantic white-sided dolphinsIlonaNo ratings yet
- Coral Remote SensingDocument15 pagesCoral Remote SensingRiskahMappileNo ratings yet
- Edited Case StudyDocument20 pagesEdited Case StudyAnonymous cLUea6DNo ratings yet
- Short Et Al - GLOBAL Seagrass DistributionDocument18 pagesShort Et Al - GLOBAL Seagrass DistributionfurkonableNo ratings yet
- Distribution and Diversity of Coastal Birds Across Habitats in Sarangani BayDocument23 pagesDistribution and Diversity of Coastal Birds Across Habitats in Sarangani Baygerman guazaNo ratings yet
- ExtinctionDocument49 pagesExtinctionShaEshaa100% (2)
- Conservation of Freshwater Bivalves at The Global Scale: Diversity, Threats and Research NeedsDocument14 pagesConservation of Freshwater Bivalves at The Global Scale: Diversity, Threats and Research NeedsAireen Diamsim MaltoNo ratings yet
- Endemic Fishes of The Gulf of CaliforniaDocument12 pagesEndemic Fishes of The Gulf of CaliforniaMarianne MtzNo ratings yet
- Bloom 2012Document15 pagesBloom 2012Fadhli LatuconsinaNo ratings yet
- Invasive Species, A Lobal ConcernDocument14 pagesInvasive Species, A Lobal ConcerneparicoNo ratings yet
- Biological Conservation: Contents Lists Available atDocument11 pagesBiological Conservation: Contents Lists Available atPhoebe Sabilla RanceNo ratings yet
- Contenido Estomacal USA FloridaDocument19 pagesContenido Estomacal USA FloridaRuth Vasquez LevyNo ratings yet
- MSU Faculty of Science and Technology student explores island biogeographyDocument5 pagesMSU Faculty of Science and Technology student explores island biogeographyYOLANDA NYARADZO MUPITANo ratings yet
- kazyak2013Document11 pageskazyak2013barti koksNo ratings yet
- Nakedgoby 2015Document8 pagesNakedgoby 2015SV PRNo ratings yet
- Articulo 14Document8 pagesArticulo 14api-286454950No ratings yet
- Do No So DuttonDocument13 pagesDo No So DuttontortugamarinaNo ratings yet
- Aquaculture Disease and Health Management Research NeedsDocument10 pagesAquaculture Disease and Health Management Research NeedsNehaNo ratings yet
- Biological Conservation: Christopher E. Cattau, Julien Martin, Wiley M. KitchensDocument8 pagesBiological Conservation: Christopher E. Cattau, Julien Martin, Wiley M. KitchensMax RamosNo ratings yet
- The Causes of Biodiversity Loss in The Marine Ecosystem and Its Effect On The WorldDocument8 pagesThe Causes of Biodiversity Loss in The Marine Ecosystem and Its Effect On The Worldapi-286948298No ratings yet
- Barracudas Supplement - Berkeley 2014Document35 pagesBarracudas Supplement - Berkeley 2014IvanakbarPurwamaskaNo ratings yet
- Velez-Rubio Et Al. 2018 - Marine DebrisDocument9 pagesVelez-Rubio Et Al. 2018 - Marine DebrisKarumbéNo ratings yet
- Convention On Biological DiversityDocument10 pagesConvention On Biological DiversityAmol ToraskarNo ratings yet
- Whit Field, 1999) Ichthyofaunal Assemblages in Estuaries - A South African Case StudyDocument36 pagesWhit Field, 1999) Ichthyofaunal Assemblages in Estuaries - A South African Case StudyAndré S. FilguerasNo ratings yet
- McCann (2014)Document11 pagesMcCann (2014)Marita AndreaNo ratings yet
- Session 3 Review River Fisheries: Ecological Basis For Management and ConservationDocument40 pagesSession 3 Review River Fisheries: Ecological Basis For Management and ConservationLove RebiraNo ratings yet
- Ecological Factors Influencing Group Sizes of River Dolphins (Inia Geoffrensis and Sotalia Fluviatilis)Document10 pagesEcological Factors Influencing Group Sizes of River Dolphins (Inia Geoffrensis and Sotalia Fluviatilis)Nataliaaa684No ratings yet
- Ecosystem Effects of Fishing in Kelp Forest Communities (Tegner, 2000)Document11 pagesEcosystem Effects of Fishing in Kelp Forest Communities (Tegner, 2000)Francisco J. OvalleNo ratings yet
- Biological Invasions As A Component of GDocument10 pagesBiological Invasions As A Component of GROHIT AGRAWALNo ratings yet
- Varit's Capstone Project PaperDocument19 pagesVarit's Capstone Project PaperVarit SuriyasomboonNo ratings yet
- Species of Concern Dusky Shark: NOAA National Marine Fisheries ServiceDocument5 pagesSpecies of Concern Dusky Shark: NOAA National Marine Fisheries ServiceJessica MarieNo ratings yet
- Cormorant Predation Linked to Declines in Walleye and Yellow Perch PopulationsDocument15 pagesCormorant Predation Linked to Declines in Walleye and Yellow Perch PopulationsDavid Orlando Carmona LagosNo ratings yet
- Biodiversity: Genetic, Species, and Habitat Diversity Ecosystem ServicesDocument92 pagesBiodiversity: Genetic, Species, and Habitat Diversity Ecosystem ServicesLena TruongNo ratings yet
- The Effects of Climate Change On Coral Reefs Evaluation EssayDocument6 pagesThe Effects of Climate Change On Coral Reefs Evaluation EssayJohnny DinhNo ratings yet
- Biol Conserv 144 1961aDocument49 pagesBiol Conserv 144 1961aArturo TejeroNo ratings yet
- Art 11Document13 pagesArt 11Jeancarlos QuispeNo ratings yet
- Threat To Biodiversity (1913101042267)Document8 pagesThreat To Biodiversity (1913101042267)Neeraj DNo ratings yet
- Braga Et Al 2012 - Feeding Ecology of Fishes An Overview of Worldwide PublicationsDocument15 pagesBraga Et Al 2012 - Feeding Ecology of Fishes An Overview of Worldwide PublicationsAdreyAntunesNo ratings yet
- LMB - Irh 87, 353-363Document11 pagesLMB - Irh 87, 353-363Lucero MorenoNo ratings yet
- 8 LRBBracken CalCOFI 2007 PDFDocument10 pages8 LRBBracken CalCOFI 2007 PDFCRNo ratings yet
- Biodiversity PDFDocument24 pagesBiodiversity PDFstaldNo ratings yet
- Fishers' Local Ecological Knowledge Indicate Migration Patterns of Tropical Freshwater Fish in An Amazonian RiverDocument19 pagesFishers' Local Ecological Knowledge Indicate Migration Patterns of Tropical Freshwater Fish in An Amazonian RiversestefanieNo ratings yet
- Conservation Status of the American Horseshoe CrabDocument41 pagesConservation Status of the American Horseshoe CrabNexi anessaNo ratings yet
- North PoleDocument5 pagesNorth PoleCarlaNo ratings yet
- Biodiversity: What is it, why is it importantDocument19 pagesBiodiversity: What is it, why is it importantManohar JadhavNo ratings yet
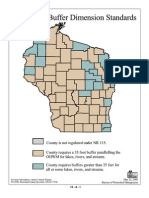
- Shoreland Vegetation - Buffer - Standards by County in WisconsinDocument29 pagesShoreland Vegetation - Buffer - Standards by County in WisconsinLori KoschnickNo ratings yet
- Regulation of Illegal Sand MiningDocument15 pagesRegulation of Illegal Sand Miningpapori35No ratings yet
- Philippine Environmental LawsDocument3 pagesPhilippine Environmental LawsTano MarkNo ratings yet
- A Genetic Classification of Floodplains: GeomorphologyDocument28 pagesA Genetic Classification of Floodplains: Geomorphologyresacoxis9No ratings yet
- Diversity and Distribution of Cyanobacteria Species in Lake Chilwa Biosphere ReserveDocument2 pagesDiversity and Distribution of Cyanobacteria Species in Lake Chilwa Biosphere ReserveIJARP PublicationsNo ratings yet
- Environmental AccountingDocument27 pagesEnvironmental AccountingFaresh Haroon100% (1)
- Analysis PWD Rate Schedule 2014Document2,144 pagesAnalysis PWD Rate Schedule 2014dxzaber80% (10)
- Plowing TheoryDocument13 pagesPlowing TheoryAnonymous 76utisSzdNo ratings yet
- Transformações de coordenadas UTM>TopográficasDocument15 pagesTransformações de coordenadas UTM>TopográficasAlEx CrUzNo ratings yet
- 13 Nu Drain Underground Drainage SystemDocument6 pages13 Nu Drain Underground Drainage SystemMahesh SavandappaNo ratings yet
- Ra 11396Document9 pagesRa 11396Ivy Galit Dula100% (1)
- Wildlife Rehabilitation Study GuideDocument8 pagesWildlife Rehabilitation Study GuideStacie BoothNo ratings yet
- 1.aronia GuideDocument148 pages1.aronia GuideDinko BećirspahićNo ratings yet
- Pimiento Et Al - MagnoliaResuinatifoliaDocument21 pagesPimiento Et Al - MagnoliaResuinatifoliamelisayalaNo ratings yet
- Facies AND Types of Hydrothermal Alteration: Alogy and Chemistry of Altered Rocks Are Not As Simple As Most of Us WouldDocument17 pagesFacies AND Types of Hydrothermal Alteration: Alogy and Chemistry of Altered Rocks Are Not As Simple As Most of Us WouldRyuzaki ChristianNo ratings yet
- Multi-sector Partnerships Key to River Rehab SuccessDocument116 pagesMulti-sector Partnerships Key to River Rehab SuccessKristia CapioNo ratings yet
- Wildlife Sanctuaries PDFDocument6 pagesWildlife Sanctuaries PDFRohit SinghNo ratings yet
- Handbook For Building Homes of EarthDocument144 pagesHandbook For Building Homes of EarthSantosh Athani100% (5)
- Study Guide On Human PopulationDocument4 pagesStudy Guide On Human Populationclaudia_614916No ratings yet
- Whittlesey's Classification of Major Agricultural RegionsDocument11 pagesWhittlesey's Classification of Major Agricultural RegionsMariam Kareem100% (4)
- Glossary GeologyDocument128 pagesGlossary GeologyMeraGurdaspurNo ratings yet
- Maine ATV MapDocument2 pagesMaine ATV MapJohn MazoNo ratings yet
- Dispute over ownership of dried creek bedDocument2 pagesDispute over ownership of dried creek bedeNo ratings yet
- Red Data Book of European Butterflies (Rhopalocera) : Nature and Environment, No. 99Document259 pagesRed Data Book of European Butterflies (Rhopalocera) : Nature and Environment, No. 99TheencyclopediaNo ratings yet
- Alemayehu Book 2006 Ethiopia Groundwater Occurrence Hydrogeological Water Quality AquiferDocument107 pagesAlemayehu Book 2006 Ethiopia Groundwater Occurrence Hydrogeological Water Quality AquiferAbiued Ejigue100% (5)
- Why Do We Need Landscaping?Document5 pagesWhy Do We Need Landscaping?ram mohan reddy100% (2)
- Treasure Hunting NewsDocument3 pagesTreasure Hunting NewsNes ConstanteNo ratings yet
- Section I: Project Description: Final Environment Assessment Report of Northern Region System Strengthening XxivDocument31 pagesSection I: Project Description: Final Environment Assessment Report of Northern Region System Strengthening XxivRajeshYadavNo ratings yet
- 03 078Document2 pages03 078Gustavo Gabriel JimenezNo ratings yet
- The Good Oil Eucalyptus Oil Distilleries in Australia PDFDocument9 pagesThe Good Oil Eucalyptus Oil Distilleries in Australia PDFMaurizio ArdizziNo ratings yet
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesFrom EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesRating: 3.5 out of 5 stars3.5/5 (56)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceFrom EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceNo ratings yet
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceFrom EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceRating: 4 out of 5 stars4/5 (632)