You might also like
- Sex Chromosome Aberrations and Transsexualism: VOL. 79, NO. 3, MARCH 2003Document2 pagesSex Chromosome Aberrations and Transsexualism: VOL. 79, NO. 3, MARCH 2003Fvg Fvg FvgNo ratings yet
- Molecular Analysis in True Hermaphrodites With Different Karyotypes and Similar PhenotypesDocument8 pagesMolecular Analysis in True Hermaphrodites With Different Karyotypes and Similar Phenotypesdanial tarikNo ratings yet
- Chromosomal Findings in Patients with Disorders of Sexual DevelopmentDocument12 pagesChromosomal Findings in Patients with Disorders of Sexual DevelopmentHorvath AlexandraNo ratings yet
- Tian 2014Document6 pagesTian 2014Lawal Bello DanchadiNo ratings yet
- Chromosome Imbalance As A Driver of Sex Disparity in DiseaseDocument12 pagesChromosome Imbalance As A Driver of Sex Disparity in DiseaseFirmanNo ratings yet
- Applied-Multiple XY-Dr. Abhay Singh YadavDocument4 pagesApplied-Multiple XY-Dr. Abhay Singh YadavImpact JournalsNo ratings yet
- Molecular Detection of Y Chromosome Microdeletions: An Irish StudyDocument6 pagesMolecular Detection of Y Chromosome Microdeletions: An Irish Studya.itatahineNo ratings yet
- Human Sex DeterminationDocument5 pagesHuman Sex DeterminationAltaicaNo ratings yet
- Forensic Science International: GeneticsDocument6 pagesForensic Science International: GeneticsIfetayo Osifeso-FakoredeNo ratings yet
- Intersex or Disorders of Sexual DevelopmentDocument19 pagesIntersex or Disorders of Sexual DevelopmentHUSAMNo ratings yet
- Identification of Sex-Specific Molecular Markers Using Restriction Site-Associated DNA SequencingDocument12 pagesIdentification of Sex-Specific Molecular Markers Using Restriction Site-Associated DNA SequencingFernando Castro EchavezNo ratings yet
- Identification of Sex-Specific Molecular Markers Using Restriction Site-Associated DNA SequencingDocument12 pagesIdentification of Sex-Specific Molecular Markers Using Restriction Site-Associated DNA SequencingBlaxez YTNo ratings yet
- NIH Public Access: Androgen Receptor Repeat Length Polymorphism Associated With Male-to-Female TranssexualismDocument9 pagesNIH Public Access: Androgen Receptor Repeat Length Polymorphism Associated With Male-to-Female TranssexualismYisus OMGNo ratings yet
- Sex Determination HumansDocument13 pagesSex Determination HumansPaweł MareckiNo ratings yet
- Abnor Alities of Hu An Sex Deter Ination: D. Tnher. Metab. DisDocument8 pagesAbnor Alities of Hu An Sex Deter Ination: D. Tnher. Metab. DisMahmoud AbouelsoudNo ratings yet
- Margarit 2000Document4 pagesMargarit 2000dad dzd adaNo ratings yet
- Handout - Sex DeterminationDocument33 pagesHandout - Sex DeterminationAyah AbusafiaNo ratings yet
- Pnas00101 0183 PDFDocument4 pagesPnas00101 0183 PDFGAMEPLAY & FUNNY MOMENTSNo ratings yet
- Y ChromosomeDocument33 pagesY Chromosomeifrahkhalid52No ratings yet
- 38591-Article Text-135841-1-10-20181015Document3 pages38591-Article Text-135841-1-10-20181015smk ypkNo ratings yet
- Sex Origin VertebratesDocument9 pagesSex Origin VertebratesBiologia MencionNo ratings yet
- Dumic 2008Document8 pagesDumic 2008Richard MacienteNo ratings yet
- XP Deletions Associated With Autism in Three FemalesDocument6 pagesXP Deletions Associated With Autism in Three FemalesRami NaNo ratings yet
- Flejter 1996Document6 pagesFlejter 1996Rahul AmbawataNo ratings yet
- Andrologia 2019Document9 pagesAndrologia 2019mayra alejandra pinilla guerreroNo ratings yet
- Emj 43 2 166 169Document4 pagesEmj 43 2 166 169Nisarga GowdaNo ratings yet
- Next-gen sequencing and karyotyping diagnose Robertsonian translocation trisomy 13Document4 pagesNext-gen sequencing and karyotyping diagnose Robertsonian translocation trisomy 13christian roblesNo ratings yet
- Burégio-Frota Et Al 2010Document5 pagesBurégio-Frota Et Al 2010dida_bispoNo ratings yet
- Isochromosome Paper-CorrectionDocument6 pagesIsochromosome Paper-CorrectionponnambalamNo ratings yet
- (L-10) - Genetics - Sep 20, 2019 - FridayDocument41 pages(L-10) - Genetics - Sep 20, 2019 - FridayOmar GillNo ratings yet
- Peart2010 PDFDocument2 pagesPeart2010 PDFMaría ElenaNo ratings yet
- Murase Et Al 2017 Genes To CellsDocument9 pagesMurase Et Al 2017 Genes To CellsVíctor Luciano Bárcena GarayNo ratings yet
- Male Cytogenetic Evaluation Prior To Assisted Reproduction Proce-Dures Performed in Mar Del Plata, ArgentinaDocument4 pagesMale Cytogenetic Evaluation Prior To Assisted Reproduction Proce-Dures Performed in Mar Del Plata, ArgentinaDevansh ShethNo ratings yet
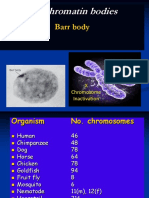
- Sex Chromatin Bodies: Barr BodyDocument45 pagesSex Chromatin Bodies: Barr BodyKhaleelMetlakNo ratings yet
- Vil Chis 2008Document6 pagesVil Chis 2008Niki MourtziNo ratings yet
- XOL-1, Primary Determinant of Sexual Fate in C. Elegans, Is A GHMP Kinase Family Member and A Structural Prototype For A Class of Developmental RegulatorsDocument14 pagesXOL-1, Primary Determinant of Sexual Fate in C. Elegans, Is A GHMP Kinase Family Member and A Structural Prototype For A Class of Developmental RegulatorsWasim ANo ratings yet
- Hum. Reprod. Update 2002 Laml 483 91Document9 pagesHum. Reprod. Update 2002 Laml 483 91Amit GoelNo ratings yet
- GRUPO 7 SX TurnerDocument6 pagesGRUPO 7 SX TurnerMariangel Suarez HenriquezNo ratings yet
- Underhill Et Al 2000Document4 pagesUnderhill Et Al 2000Ava GodhardtNo ratings yet
- Hum. Reprod.-2008-Bellver-278-84Document7 pagesHum. Reprod.-2008-Bellver-278-84Grigoriu RaduNo ratings yet
- Swyer Syndrome 5 7 14Document20 pagesSwyer Syndrome 5 7 14ireneaureliaNo ratings yet
- Chromosome Rearrangements and Survival of AndrogenDocument9 pagesChromosome Rearrangements and Survival of AndrogenJose Franco NaviaNo ratings yet
- Polygenic Sex Determination: Quick GuideDocument3 pagesPolygenic Sex Determination: Quick GuideSohan kunduNo ratings yet
- Human Y Chromosome, Sex Determination, and Spermatogenesis - A Feminist ViewDocument10 pagesHuman Y Chromosome, Sex Determination, and Spermatogenesis - A Feminist ViewIsah Enemaku JonathanNo ratings yet
- NANTHINIDocument28 pagesNANTHINIADHITHIYA VNo ratings yet
- Gender Differences in Neurodevelopmental Disorders Support "Female Protective ModelDocument11 pagesGender Differences in Neurodevelopmental Disorders Support "Female Protective Modelcinfer75No ratings yet
- Mendelian Inheritance in Man: Catalogs of Autosomal Dominant, Autosomal Recessive, and X-Linked PhenotypesFrom EverandMendelian Inheritance in Man: Catalogs of Autosomal Dominant, Autosomal Recessive, and X-Linked PhenotypesNo ratings yet
- Selectivity of Hyaluronic Acid Binding For Spermatozoa With Normal Tygerberg Strict MorphologyDocument7 pagesSelectivity of Hyaluronic Acid Binding For Spermatozoa With Normal Tygerberg Strict MorphologyMafe HerreraNo ratings yet
- Bongers 1999 Viable Androgenetic Yy Genotypes ofDocument4 pagesBongers 1999 Viable Androgenetic Yy Genotypes ofAndreea ȘerbanNo ratings yet
- Genetically Homozygous Choriocarcinoma Following Hydatidiform MoleDocument5 pagesGenetically Homozygous Choriocarcinoma Following Hydatidiform MoleFeby HidasariNo ratings yet
- 575854100sex-Linked InheritanceDocument18 pages575854100sex-Linked InheritanceMohammad Faizan RazaNo ratings yet
- Autosomal Deletion SyndromesDocument50 pagesAutosomal Deletion SyndromesZainab Jamal SiddiquiNo ratings yet
- 第三期 北医三院 MarsalaDocument16 pages第三期 北医三院 MarsalazjuwindNo ratings yet
- Genetic Counseling in TranslocationsDocument15 pagesGenetic Counseling in TranslocationsAsma HMILANo ratings yet
- 47, XYY Syndrome and Its Association With Male Infertility: CaseDocument3 pages47, XYY Syndrome and Its Association With Male Infertility: CaseprasadbheemNo ratings yet
- Calafell 1999Document10 pagesCalafell 1999friiday.qNo ratings yet
- Low Rates of Pregnancy Termination For Prenatally Diagnosed Klinefelter Syndrome and Other Sex Chromosome PolysomiesDocument5 pagesLow Rates of Pregnancy Termination For Prenatally Diagnosed Klinefelter Syndrome and Other Sex Chromosome PolysomiesAltaicaNo ratings yet
- Pediatric Disorders of Sex DevelopmentDocument3 pagesPediatric Disorders of Sex DevelopmentMuhammad Amri KautsarNo ratings yet
- Xy Sex Determination SystemDocument7 pagesXy Sex Determination Systemafricano cregNo ratings yet
- Jurnal Reading11 2 Forensik PDFDocument5 pagesJurnal Reading11 2 Forensik PDFrennyNo ratings yet
- Pkol Wvs 732 1915 l500 Kuhp EnglishDocument117 pagesPkol Wvs 732 1915 l500 Kuhp EnglishJody DewantoroNo ratings yet
- Body Part Reconstruction in Mutilated Body Identification CaseDocument8 pagesBody Part Reconstruction in Mutilated Body Identification CaserennyNo ratings yet
- Pkol Wvs 732 1915 l500 Kuhp EnglishDocument117 pagesPkol Wvs 732 1915 l500 Kuhp EnglishJody DewantoroNo ratings yet
- Template Artikel - EnglishDocument1 pageTemplate Artikel - EnglishAkmal MuhniNo ratings yet
- Pkol Wvs 732 1915 l500 Kuhp EnglishDocument117 pagesPkol Wvs 732 1915 l500 Kuhp EnglishJody DewantoroNo ratings yet
- Development and Targeted Application of A Rapid QF-PCR Test For Sex Chromosome ImbalanceDocument10 pagesDevelopment and Targeted Application of A Rapid QF-PCR Test For Sex Chromosome ImbalancerennyNo ratings yet
- Asfiksia DeathDocument13 pagesAsfiksia DeathrennyNo ratings yet
- Fate of SRY, PABY, DYS1, DYZ3 and DYZ1 loci in sex chromosomal anomalies patientsDocument11 pagesFate of SRY, PABY, DYS1, DYZ3 and DYZ1 loci in sex chromosomal anomalies patientsrenny100% (1)
- Jurnal LukatembakDocument10 pagesJurnal LukatembakrennyNo ratings yet
- Summary Fatty Alcohols LFRMIDocument2 pagesSummary Fatty Alcohols LFRMILuis Fernando Ramos Méndez IrisNo ratings yet
- Genetic DiversityDocument31 pagesGenetic DiversityAbegail AvilaNo ratings yet
- Dapuss 6Document5 pagesDapuss 6SasmitaNo ratings yet
- Lesson PlanDocument2 pagesLesson PlanQueenie O Cerlos100% (2)
- Class 9 The Fundamental Unit of Life Assignment 7Document3 pagesClass 9 The Fundamental Unit of Life Assignment 7Madhusudan BanerjeeNo ratings yet
- Lab Investigation AP 3 - Constructing Evol Lineages With DNA BLASTDocument7 pagesLab Investigation AP 3 - Constructing Evol Lineages With DNA BLASTKeri Gobin SamarooNo ratings yet
- Order 157489 - PSC Requisition - Ulta Lab TestsDocument2 pagesOrder 157489 - PSC Requisition - Ulta Lab Testszarrin77No ratings yet
- Xu2015 Article RapidGenerationAndAnalysisOfExDocument13 pagesXu2015 Article RapidGenerationAndAnalysisOfExIrsyadKamalNo ratings yet
- Trees Within Trees: Genes and Species, Molecules and MorphologyDocument17 pagesTrees Within Trees: Genes and Species, Molecules and MorphologyWendimu GirmaNo ratings yet
- gr11 Mitosis Meiosis QuizDocument3 pagesgr11 Mitosis Meiosis Quizapi-250802146No ratings yet
- Organic Evolution TheoriesDocument8 pagesOrganic Evolution TheoriesCynthia Adeline SNo ratings yet
- IJB Vol 14 No 1 P 533 542Document10 pagesIJB Vol 14 No 1 P 533 542Muhammad Amir MaqboolNo ratings yet
- NCBIDocument13 pagesNCBIMuaris BowoNo ratings yet
- Cambridge A and AS Biology - Overview of Exam Questions and Worked ExamplesDocument3 pagesCambridge A and AS Biology - Overview of Exam Questions and Worked ExamplesqhlwjuvrvqygwglzegNo ratings yet
- V1-3 DNA WiSe2324Document52 pagesV1-3 DNA WiSe2324Waheed AhmadNo ratings yet
- Molecular Biology Lab - Report تقرير عن معمل بيولوجيا جزيئيةDocument19 pagesMolecular Biology Lab - Report تقرير عن معمل بيولوجيا جزيئيةManal AL-KhouliNo ratings yet
- trnH-psbA Plant DNA BarcodeDocument7 pagestrnH-psbA Plant DNA BarcoderaunakmsNo ratings yet
- Rani Anak Mat Case 4 ReportDocument5 pagesRani Anak Mat Case 4 ReportRani anak matNo ratings yet
- Rajkumar Biology Unit - 9 by RajatDocument12 pagesRajkumar Biology Unit - 9 by RajatprasannandaNo ratings yet
- Plant Genetic Resources and ConservationDocument10 pagesPlant Genetic Resources and ConservationMahathir Mohmed100% (32)
- Test Bank For Larsens Human Embryology 5th Edition Gary Schoenwolf Steven Bleyl Philip Brauer Philippa Francis WestDocument3 pagesTest Bank For Larsens Human Embryology 5th Edition Gary Schoenwolf Steven Bleyl Philip Brauer Philippa Francis WestPauline Chavez100% (31)
- Mitosis: J. Richard McintoshDocument17 pagesMitosis: J. Richard McintoshAlejandro CastañedaNo ratings yet
- Cell我们毕业啦 - Cell Differentiation and Key FactorsDocument46 pagesCell我们毕业啦 - Cell Differentiation and Key Factorsthri zonartiNo ratings yet
- Revision - Why Do We Fall Ill NotesDocument15 pagesRevision - Why Do We Fall Ill NotesAtharva VarshneyNo ratings yet
- (Cancer Drug Discovery and Development) A. Ganesan (Auth.), Frank E. Koehn (Eds.) - Natural Products and Cancer Drug Discovery-Springer-Verlag New York (2013)Document248 pages(Cancer Drug Discovery and Development) A. Ganesan (Auth.), Frank E. Koehn (Eds.) - Natural Products and Cancer Drug Discovery-Springer-Verlag New York (2013)shuvvro dhaNo ratings yet
- List of Pharmaceutical Companies in India With Address - Pharma CompaniesDocument73 pagesList of Pharmaceutical Companies in India With Address - Pharma Companiessrivin naik58% (12)
- Petition Response Letter From FDA CDER To Insys Therapeutics, IncDocument12 pagesPetition Response Letter From FDA CDER To Insys Therapeutics, IncKyleJaegerNo ratings yet
- In Situ Hybridization PDFDocument5 pagesIn Situ Hybridization PDFAnonymous ThXYCGkLCLNo ratings yet
- Gene Cloning TechnologyDocument16 pagesGene Cloning TechnologyNguyen Thanh Anh NiiroNo ratings yet
- Planaria Lab ReportDocument3 pagesPlanaria Lab Reportapi-194648534No ratings yet