You might also like
- CARISMA - Cherry Tomato - en PDFDocument4 pagesCARISMA - Cherry Tomato - en PDFRica BurnichiNo ratings yet
- Lycopene in Tomatoes: Chemical and Physical Properties Affected by Food ProcessingDocument43 pagesLycopene in Tomatoes: Chemical and Physical Properties Affected by Food Processingnur halimaNo ratings yet
- How to Make Tomato JamDocument4 pagesHow to Make Tomato JamLisly DizonNo ratings yet
- Soil Health, Soil Biology, Soilborne Diseases and Sustainable Agriculture: A GuideFrom EverandSoil Health, Soil Biology, Soilborne Diseases and Sustainable Agriculture: A GuideRating: 5 out of 5 stars5/5 (1)
- Tomatoes and Tomato Products - Nutritional, Medicinal and Therapeutic PropertiesDocument664 pagesTomatoes and Tomato Products - Nutritional, Medicinal and Therapeutic PropertiesAndreea MerţNo ratings yet
- Citrus Fruit: Biology, Technology and EvaluationFrom EverandCitrus Fruit: Biology, Technology and EvaluationRating: 3.5 out of 5 stars3.5/5 (2)
- Chapter 1 DoneDocument9 pagesChapter 1 DoneLemuel L. Madridejos0% (2)
- ORGANIC VEGETABLE GARDENING: How to Grow Your Vegetables and Start a Healthy Garden at Home. A Step-by-Step Guide for Beginners (2022)From EverandORGANIC VEGETABLE GARDENING: How to Grow Your Vegetables and Start a Healthy Garden at Home. A Step-by-Step Guide for Beginners (2022)No ratings yet
- Nutritional Value of Cherry Tomatoes (Lycopersicon Esculentum Cv. Naomi F1) Harvested at Different Ripening StagesDocument7 pagesNutritional Value of Cherry Tomatoes (Lycopersicon Esculentum Cv. Naomi F1) Harvested at Different Ripening Stagesgonzalo_hbNo ratings yet
- F & V Lecture #5 & 6 General Properties of Fruits and VegetablesDocument5 pagesF & V Lecture #5 & 6 General Properties of Fruits and VegetablesZohaib ArshadNo ratings yet
- Fruit and Vegetables: Your Dietary Essentials: The International Year of Fruits and Vegetables, 2021, Background PaperFrom EverandFruit and Vegetables: Your Dietary Essentials: The International Year of Fruits and Vegetables, 2021, Background PaperNo ratings yet
- Effects of Red Tomatoes As A Ripening Accelerator On Mature Green TomatoesDocument4 pagesEffects of Red Tomatoes As A Ripening Accelerator On Mature Green TomatoesKelly EstilongNo ratings yet
- Ssac 243Document19 pagesSsac 243Fake HankNo ratings yet
- TomatoesDocument6 pagesTomatoesNataly Cruz VenturinaNo ratings yet
- Characterization of Sugars and Organic Acids in Commercial Varieties of Table GrapesDocument7 pagesCharacterization of Sugars and Organic Acids in Commercial Varieties of Table GrapesHala LalaNo ratings yet
- Tomato Post-Harvest Physiology As A VegetableDocument14 pagesTomato Post-Harvest Physiology As A Vegetablechristom karungariNo ratings yet
- 1 IntroductionDocument4 pages1 IntroductionPritika RaiNo ratings yet
- Bitter Melon Bioactive Composition and Health Benefits A Review - Tan Et Al. 2015Document23 pagesBitter Melon Bioactive Composition and Health Benefits A Review - Tan Et Al. 2015Fatima HerreraNo ratings yet
- Crop Protection ProposalDocument9 pagesCrop Protection Proposalbart mitch CutamoraNo ratings yet
- Research 1Document5 pagesResearch 1Guene BinwagNo ratings yet
- Physical and Chemical Properties of Fruits and VegetableDocument4 pagesPhysical and Chemical Properties of Fruits and VegetableStatio NeryNo ratings yet
- Citrus LimonoidsDocument47 pagesCitrus LimonoidsMarie Astrid AmboyNo ratings yet
- The Physiochemical and Microbiological Quality of Fresh Tomatoes Juice and Tomatoes Paste Sold in Abraka MarketDocument10 pagesThe Physiochemical and Microbiological Quality of Fresh Tomatoes Juice and Tomatoes Paste Sold in Abraka MarketInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Analysis of Tomato Post-Harvest Properties Fruit CDocument17 pagesAnalysis of Tomato Post-Harvest Properties Fruit CMădălina ȘtefanNo ratings yet
- Introduction To Fruits and VegetableDocument48 pagesIntroduction To Fruits and VegetableKoushik DasNo ratings yet
- Tomato Jam Processing GuideDocument4 pagesTomato Jam Processing GuideCandy Concepcion PadizNo ratings yet
- Tugas Bahasa InggrisDocument7 pagesTugas Bahasa Inggriselisa gobelNo ratings yet
- Post HarvestDocument119 pagesPost HarvestTrung Le67% (3)
- Horticulturae 09 00163 v2Document13 pagesHorticulturae 09 00163 v2Eric Arturo Medina SuNo ratings yet
- Tomato CanningDocument16 pagesTomato CanningMUHAMMAD UMARNo ratings yet
- Introduction To Fruits and Vegetables 1122Document71 pagesIntroduction To Fruits and Vegetables 1122Dr-Uadal SinghNo ratings yet
- Caleb PROJECT ProposalDocument21 pagesCaleb PROJECT ProposalthelonewolfmerchNo ratings yet
- Molecules 25 05604Document51 pagesMolecules 25 05604Ivana TomazNo ratings yet
- FST - 223Document92 pagesFST - 223Karthikeyan BalakrishnanNo ratings yet
- Genetic and Environmental Factors Affect Tomato Flesh ColorDocument6 pagesGenetic and Environmental Factors Affect Tomato Flesh ColorAlex SullivanNo ratings yet
- Characterization of Phenolics and Sugars in Melicoccus bijugatus Fruits by LC-UV/Vis, LC-MS/MS and GCDocument8 pagesCharacterization of Phenolics and Sugars in Melicoccus bijugatus Fruits by LC-UV/Vis, LC-MS/MS and GCnatale_carvalhoNo ratings yet
- SynopsisDocument32 pagesSynopsisFerez DedioNo ratings yet
- Distribution and inheritance of aroma compounds in tangerine hybridsDocument12 pagesDistribution and inheritance of aroma compounds in tangerine hybridsnishi@sainiNo ratings yet
- Science Investigatory Project Father John Karash Memorial High SchoolDocument12 pagesScience Investigatory Project Father John Karash Memorial High Schoolxylene cuaresmaNo ratings yet
- Harvey Lacson Research 2Document28 pagesHarvey Lacson Research 2Eltanal KenNo ratings yet
- 8 Nataša RistovskaDocument5 pages8 Nataša RistovskaAnggreini FajarNo ratings yet
- Lychee FlavourDocument58 pagesLychee FlavourshishirchemNo ratings yet
- HannahDocument1 pageHannahFrances PaulineNo ratings yet
- Santoso (5.81) .Bab IDocument13 pagesSantoso (5.81) .Bab Isharktank4youNo ratings yet
- Lomato FruitDocument3 pagesLomato FruitROSE TASINo ratings yet
- Fruit and Vegetable Process Tecnology Chapter 3Document89 pagesFruit and Vegetable Process Tecnology Chapter 3robel kassawNo ratings yet
- Artirulo Ref Licopeno 6Document7 pagesArtirulo Ref Licopeno 6Eric Arturo Medina SuNo ratings yet
- Sample Research Proposal (8).1687444473233Document13 pagesSample Research Proposal (8).1687444473233vricodamianfrancelyrNo ratings yet
- Artikel Nadila B InggrisDocument6 pagesArtikel Nadila B InggrisAtika Rahma AyuNo ratings yet
- Strawberry FlavorDocument5 pagesStrawberry FlavorBrían Goh100% (2)
- Analysis of Phenolic Compounds by HPLC and LC-MSDocument9 pagesAnalysis of Phenolic Compounds by HPLC and LC-MSBrenda D'AcunhaNo ratings yet
- Vegetable Juice Analysis Reveals Key NutrientsDocument18 pagesVegetable Juice Analysis Reveals Key NutrientsMark Jacob FelicianoNo ratings yet
- Jurnal Teknologi Dan Industri Pertanian Indonesia: KeywordsDocument5 pagesJurnal Teknologi Dan Industri Pertanian Indonesia: KeywordsYusdike AdystiaNo ratings yet
- FOOD AND NUTRITIONAL ANALYSIS Fruits and Fruit Products-334-341Document8 pagesFOOD AND NUTRITIONAL ANALYSIS Fruits and Fruit Products-334-341fernandoferozNo ratings yet
- Subject-Chemistry Investigatory ProjectDocument21 pagesSubject-Chemistry Investigatory Projectarjun b rNo ratings yet
- Processing, Handling and Storage of Agricultural Product 2Document6 pagesProcessing, Handling and Storage of Agricultural Product 2LittleagleNo ratings yet
- 9 LCA CertificationDocument1 page9 LCA CertificationAnouer BelkadiNo ratings yet
- 10 AutodeskDocument1 page10 AutodeskAnouer BelkadiNo ratings yet
- 7-Anouar Belkadi LED ProfissionalDocument1 page7-Anouar Belkadi LED ProfissionalAnouer BelkadiNo ratings yet
- 10 AutodeskDocument1 page10 AutodeskAnouer BelkadiNo ratings yet
- 5-Anouar Belkadi Led PasseportDocument1 page5-Anouar Belkadi Led PasseportAnouer BelkadiNo ratings yet
- 7-Anouar Belkadi LED ProfissionalDocument1 page7-Anouar Belkadi LED ProfissionalAnouer BelkadiNo ratings yet
- 8-Zumtobel Lighting ControlDocument1 page8-Zumtobel Lighting ControlAnouer BelkadiNo ratings yet
- 3-The Official EF SET Certificate™ - Anouar BelkadiDocument3 pages3-The Official EF SET Certificate™ - Anouar BelkadiAnouer BelkadiNo ratings yet
- 6-Anouar Belkadi Associate Certified PhilipsDocument1 page6-Anouar Belkadi Associate Certified PhilipsAnouer BelkadiNo ratings yet
- 6-Anouar Belkadi Associate Certified PhilipsDocument1 page6-Anouar Belkadi Associate Certified PhilipsAnouer BelkadiNo ratings yet
- 5-Anouar Belkadi LED PASSEPORTDocument1 page5-Anouar Belkadi LED PASSEPORTAnouer BelkadiNo ratings yet
- File 000008Document1 pageFile 000008Anouer BelkadiNo ratings yet
- LicenseDocument1 pageLicenseElvin EbinNo ratings yet
- 9-LCA CertificationDocument1 page9-LCA CertificationAnouer BelkadiNo ratings yet
- Dialux CertificationDocument1 pageDialux CertificationAnouer Belkadi100% (1)
- IP&IK TestDocument5 pagesIP&IK TestAnouer BelkadiNo ratings yet
- DIALux Partner CertificateDocument1 pageDIALux Partner CertificateAnouer BelkadiNo ratings yet
- File 000034Document5 pagesFile 000034Anouer BelkadiNo ratings yet
- Test Report IEC 62493 Luminaires Assessment of Lighting Equipment Related To Human Exposure To Electromagnetic FieldsDocument6 pagesTest Report IEC 62493 Luminaires Assessment of Lighting Equipment Related To Human Exposure To Electromagnetic FieldsAnouer BelkadiNo ratings yet
- Aging Test Report (3000hours) LM84Document9 pagesAging Test Report (3000hours) LM84Anouer Belkadi100% (1)
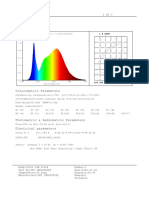
- Colorimetric Parameters: 1.0 6.191e+001mw/nm 1.2 SpectrumDocument1 pageColorimetric Parameters: 1.0 6.191e+001mw/nm 1.2 SpectrumAnouer BelkadiNo ratings yet
- MathType Commands 6 For Word1Document1 pageMathType Commands 6 For Word1thanhxuan_vn14No ratings yet
- Shenzhen Southern LCS Compliance Testing Laboratory LTD.: Streetlight Photometric Test ReportDocument10 pagesShenzhen Southern LCS Compliance Testing Laboratory LTD.: Streetlight Photometric Test ReportAnouer BelkadiNo ratings yet
- Annexe Acréditation CnasDocument173 pagesAnnexe Acréditation CnasAnouer BelkadiNo ratings yet
- Greenhouse Energy Study Optimises Shape & MaterialsDocument25 pagesGreenhouse Energy Study Optimises Shape & MaterialsAnouer BelkadiNo ratings yet
- Scholarship Cover Letter GuidlinesDocument12 pagesScholarship Cover Letter GuidlinesJacquelineLynneNo ratings yet
- Middle Power LED Series 3030: Features & BenefitsDocument25 pagesMiddle Power LED Series 3030: Features & BenefitsAnouer BelkadiNo ratings yet
- DEKRA - Application / Order Form: Application To Perform The Services Selected in This FormDocument2 pagesDEKRA - Application / Order Form: Application To Perform The Services Selected in This FormAnouer BelkadiNo ratings yet
- Test Report LM80Document13 pagesTest Report LM80Anouer BelkadiNo ratings yet
- Sample Paper 2Document7 pagesSample Paper 2ranitkumar14No ratings yet
- Estrogens, ConjugatedDocument10 pagesEstrogens, ConjugatedJuan PerezNo ratings yet
- EDTA METHOD (Sulphate)Document2 pagesEDTA METHOD (Sulphate)Bhinitha Chandrasagaran100% (1)
- More Efficient Synthesis of Alkoxybenzazepines IntermediateDocument5 pagesMore Efficient Synthesis of Alkoxybenzazepines IntermediateDipti DodiyaNo ratings yet
- New QB Acid and Base 3Document28 pagesNew QB Acid and Base 3Irmak CoşkunNo ratings yet
- 2017-Carbon Nano-Onions - Unique Carbon Nanostructures With Fascinating Properties and Their Potential ApplicationsDocument18 pages2017-Carbon Nano-Onions - Unique Carbon Nanostructures With Fascinating Properties and Their Potential Applicationsparra MedinaNo ratings yet
- Gamma-HB SynthesisDocument8 pagesGamma-HB Synthesisedman9864No ratings yet
- Coordination Compounds Multiple C. QuesDocument6 pagesCoordination Compounds Multiple C. QuesShivam KumarNo ratings yet
- Series 4100 Translucent Calendered Film: Applications & FeaturesDocument2 pagesSeries 4100 Translucent Calendered Film: Applications & Featuresgerardo_medina_92No ratings yet
- Carbon NanotubesDocument26 pagesCarbon NanotubesTanuj JoshiNo ratings yet
- Disinfection of Dental ImpressionsDocument4 pagesDisinfection of Dental ImpressionsswathiNo ratings yet
- Nitrogen FixationDocument19 pagesNitrogen FixationSidharth0% (1)
- 8 - Photolithography Part 2 and Other LithographyDocument39 pages8 - Photolithography Part 2 and Other LithographyALI HAIDERNo ratings yet
- Topic 1 - Cell Biology (Bioninja Summaries)Document10 pagesTopic 1 - Cell Biology (Bioninja Summaries)rachelNo ratings yet
- Hyperchain X Official WhitepaperDocument5 pagesHyperchain X Official Whitepaperunreal2No ratings yet
- Effect of Roasting On LactonesDocument9 pagesEffect of Roasting On LactonesRahma YuliaNo ratings yet
- Gold Plating: Alan BlairDocument2 pagesGold Plating: Alan BlairHiden HidenNo ratings yet
- Electroplating Test InterpretationDocument3 pagesElectroplating Test InterpretationR.SubramanianNo ratings yet
- Exploring Thermochromic Behaviour of Hydrated Hybrid Perovskites in Solar CellDocument5 pagesExploring Thermochromic Behaviour of Hydrated Hybrid Perovskites in Solar Cellatulchaudhari84No ratings yet
- Antoine Coefficients For Vapor PressureDocument32 pagesAntoine Coefficients For Vapor PressureJesicaidat9No ratings yet
- General Familiarity With Other NDT Methods Module 4Document4 pagesGeneral Familiarity With Other NDT Methods Module 4Weld Bro SandeepNo ratings yet
- AMAZONITE- THE GREEN VARIETY OF MICROCLINEDocument325 pagesAMAZONITE- THE GREEN VARIETY OF MICROCLINEGabriel Teixeira0% (1)
- Chapter 21Document15 pagesChapter 21Chandra ParkNo ratings yet
- The Study of A Synergetic Effect OverDocument5 pagesThe Study of A Synergetic Effect OverPascale PaulNo ratings yet
- Cellular Signal Transduction: The Biochemistry and Molecular Biology Department of CMUDocument98 pagesCellular Signal Transduction: The Biochemistry and Molecular Biology Department of CMUKrisda JitrakbumrungNo ratings yet
- Substitution Elimination FlowchartDocument2 pagesSubstitution Elimination FlowchartAyah Al-AnaniNo ratings yet
- at Urinary CalculiDocument2 pagesat Urinary Calculigaratrombozier100% (1)
- A Detailed Lesson Plan in Science 9iDocument7 pagesA Detailed Lesson Plan in Science 9iBaby Joy Jovita-BasubasNo ratings yet
- Tissue Staining Methods and TechniquesDocument21 pagesTissue Staining Methods and TechniquesBhavna RamkissoonNo ratings yet
- Bio 2Document3 pagesBio 2ganchimeg gankhuuNo ratings yet
- The Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsFrom EverandThe Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsRating: 4 out of 5 stars4/5 (146)
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincFrom EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincRating: 3.5 out of 5 stars3.5/5 (137)
- The Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableFrom EverandThe Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableRating: 3.5 out of 5 stars3.5/5 (22)
- Meltdown: Nuclear disaster and the human cost of going criticalFrom EverandMeltdown: Nuclear disaster and the human cost of going criticalRating: 5 out of 5 stars5/5 (5)
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolFrom EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolNo ratings yet
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeFrom EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeRating: 4.5 out of 5 stars4.5/5 (3)
- Handbook of Formulating Dermal Applications: A Definitive Practical GuideFrom EverandHandbook of Formulating Dermal Applications: A Definitive Practical GuideNo ratings yet
- The Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookFrom EverandThe Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookNo ratings yet
- Essential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilFrom EverandEssential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilRating: 5 out of 5 stars5/5 (1)
- Coating and Drying Defects: Troubleshooting Operating ProblemsFrom EverandCoating and Drying Defects: Troubleshooting Operating ProblemsRating: 5 out of 5 stars5/5 (1)
- Chemistry: a QuickStudy Laminated Reference GuideFrom EverandChemistry: a QuickStudy Laminated Reference GuideRating: 5 out of 5 stars5/5 (1)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeFrom EverandChemistry for Breakfast: The Amazing Science of Everyday LifeRating: 4.5 out of 5 stars4.5/5 (90)
- Monkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeFrom EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeRating: 4 out of 5 stars4/5 (1)
- Formulating, Packaging, and Marketing of Natural Cosmetic ProductsFrom EverandFormulating, Packaging, and Marketing of Natural Cosmetic ProductsNo ratings yet
- The Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsFrom EverandThe Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsRating: 5 out of 5 stars5/5 (3)
- Science Goes Viral: Captivating Accounts of Science in Everyday LifeFrom EverandScience Goes Viral: Captivating Accounts of Science in Everyday LifeRating: 5 out of 5 stars5/5 (1)
- Chemistry: 1001 Practice Problems For Dummies (+ Free Online Practice)From EverandChemistry: 1001 Practice Problems For Dummies (+ Free Online Practice)No ratings yet
- Transformer: The Deep Chemistry of Life and DeathFrom EverandTransformer: The Deep Chemistry of Life and DeathRating: 4.5 out of 5 stars4.5/5 (13)
- AP® Chemistry Crash Course, For the 2020 Exam, Book + Online: Get a Higher Score in Less TimeFrom EverandAP® Chemistry Crash Course, For the 2020 Exam, Book + Online: Get a Higher Score in Less TimeRating: 5 out of 5 stars5/5 (1)
- A Perfect Red: Empire, Espionage, and the Quest for the Color of DesireFrom EverandA Perfect Red: Empire, Espionage, and the Quest for the Color of DesireRating: 4 out of 5 stars4/5 (129)