You might also like
- Devlin Biochemistry With Clinical CorrelationsDocument1,241 pagesDevlin Biochemistry With Clinical CorrelationsJorge100% (5)
- Summary of Georgia Immunization Requirements For Child Care & School AttendanceDocument1 pageSummary of Georgia Immunization Requirements For Child Care & School AttendanceWSB-TV Assignment DeskNo ratings yet
- 3753 Full PDFDocument3 pages3753 Full PDFJames WatsonNo ratings yet
- Foundations in Microbiology Basic Principles 10th Edition Talaro Test BankDocument35 pagesFoundations in Microbiology Basic Principles 10th Edition Talaro Test Bankhieudermotjm7w100% (28)
- Paper I PDFDocument6 pagesPaper I PDFErick FariasNo ratings yet
- Benazet El Al. Science (2009) A Self-Regulatory System of Interlinked SignalingDocument5 pagesBenazet El Al. Science (2009) A Self-Regulatory System of Interlinked SignalingnovrodNo ratings yet
- Science 1213229Document5 pagesScience 12132291592162022No ratings yet
- Ultrahigh Porosity in Metal-Organic Frameworks: 'KeeffeDocument6 pagesUltrahigh Porosity in Metal-Organic Frameworks: 'Keeffeクマール ヴァンツNo ratings yet
- Singer Et Al2004Document6 pagesSinger Et Al2004Firuza RahimovaNo ratings yet
- A Physical Map of 30,000 Human Genes: J. Am. Chem. Soc. 117, 4193 (1995) ), As DemonstratDocument3 pagesA Physical Map of 30,000 Human Genes: J. Am. Chem. Soc. 117, 4193 (1995) ), As DemonstratRahmat Eko SanjayaNo ratings yet
- Gregg 2010Document5 pagesGregg 2010Daniel Nuno Vancetto BorgesNo ratings yet
- PWYW1Document3 pagesPWYW1Andrew GarfieldNo ratings yet
- Boletin de Arquelologia PUCP No. 16 (2014) Número 16. Los Rostros de Huari Perspectivas Interregionales Sobre El Horizonte MedioDocument5 pagesBoletin de Arquelologia PUCP No. 16 (2014) Número 16. Los Rostros de Huari Perspectivas Interregionales Sobre El Horizonte MedioPareja Anyosa DanteNo ratings yet
- Judy MikovitsDocument7 pagesJudy MikovitsJ Elver SilvaNo ratings yet
- PankjoyDocument2 pagesPankjoyMonitchelle Cristina do NascimentoNo ratings yet
- Nuclei Get TAN LinesDocument3 pagesNuclei Get TAN LinesArpan NandyNo ratings yet
- Excitation of Orbital Angular Momentum Resonances in Helically Twisted Photonic Crystal FiberDocument4 pagesExcitation of Orbital Angular Momentum Resonances in Helically Twisted Photonic Crystal Fiberthillai saravananNo ratings yet
- Magnetic Skyrmions: Advances in Physics and Potential ApplicationsDocument5 pagesMagnetic Skyrmions: Advances in Physics and Potential ApplicationsAgtc TandayNo ratings yet
- Reconstructing signaling networks from single-cell dataDocument8 pagesReconstructing signaling networks from single-cell dataarchsarkNo ratings yet
- E.Behnke Et Al., Science, 319, (2008) - Spin Dependent WIMP Limits From A Bubble ChamberDocument4 pagesE.Behnke Et Al., Science, 319, (2008) - Spin Dependent WIMP Limits From A Bubble ChamberIvan FelisNo ratings yet
- 4D Electron Tomography ScienceDocument6 pages4D Electron Tomography Sciencehappynewyear11No ratings yet
- Teleport Nonclassical WavepacklightDocument4 pagesTeleport Nonclassical Wavepacklighth tytionNo ratings yet
- Simulating Dynamical Features of Escape Panic: Dirk Helbing, Illeâs Farkas & Tamaâs VicsekDocument4 pagesSimulating Dynamical Features of Escape Panic: Dirk Helbing, Illeâs Farkas & Tamaâs VicsektariqNo ratings yet
- OSTROM, Elinor. A General Framework For Analizing Sustaintability of Socio Ecological SystemsDocument5 pagesOSTROM, Elinor. A General Framework For Analizing Sustaintability of Socio Ecological SystemsLizbeth Diaz RedolfoNo ratings yet
- Parrenin Science 2013Document5 pagesParrenin Science 2013Camilo CastellanosNo ratings yet
- Quantom Dot Induced Phase Stabilization of the Cspbi3 Perovskite for High Efficiency PhotovoltaicsDocument5 pagesQuantom Dot Induced Phase Stabilization of the Cspbi3 Perovskite for High Efficiency PhotovoltaicsabuhurairabscNo ratings yet
- The Following Resources Related To This Article Are Available Online atDocument5 pagesThe Following Resources Related To This Article Are Available Online atihzaoloanNo ratings yet
- Active Normal Faulting in The Upper Rhine Graben and Paleoseismic Identification of The 1356 Basel Earthquake.Document5 pagesActive Normal Faulting in The Upper Rhine Graben and Paleoseismic Identification of The 1356 Basel Earthquake.marceloahumada1983No ratings yet
- 5-2013-Research-Multiplex Genome Engineering Using CRISPRCas SystemsDocument6 pages5-2013-Research-Multiplex Genome Engineering Using CRISPRCas SystemsCristian Felipe Sandoval QuiñonezNo ratings yet
- Ross 2013Document5 pagesRoss 2013zune153No ratings yet
- T. Rex Was A Slow RunnerDocument4 pagesT. Rex Was A Slow RunnerozzaibNo ratings yet
- Barrangou 20 Et 20 Al 202007Document6 pagesBarrangou 20 Et 20 Al 202007ירדן לויןNo ratings yet
- In Modern Shells Southern Nevada: Taylor AstrophysDocument4 pagesIn Modern Shells Southern Nevada: Taylor AstrophysVitaliy T.No ratings yet
- Suppression of Rain and Snow by Urban and Industrial Air PollutionDocument6 pagesSuppression of Rain and Snow by Urban and Industrial Air PollutionDavidNo ratings yet
- Science (New York NY) 2003 TemelesDocument4 pagesScience (New York NY) 2003 TemelesmariaNo ratings yet
- Articulo Cientico 2Document5 pagesArticulo Cientico 2Elvis AndíaNo ratings yet
- A Logic-Gated Nanorobot For Targeted Transport of Molecular PayloadsDocument5 pagesA Logic-Gated Nanorobot For Targeted Transport of Molecular PayloadsJohnathonNo ratings yet
- Chen and Tung 2014 ScienceDocument8 pagesChen and Tung 2014 Science1739598671No ratings yet
- Acfrogcfqtzu2xnftwqjnegs7ygzf1guo4 K Hves Qm5urgs6j7bracmbcs4px1djm2jtjpktog2kwjhdexqbirxrvocnrlkvl6dixeprgtv0xlrmwujhdy5qi6md71yupr4hwrgyj9ym1ev A6Document6 pagesAcfrogcfqtzu2xnftwqjnegs7ygzf1guo4 K Hves Qm5urgs6j7bracmbcs4px1djm2jtjpktog2kwjhdexqbirxrvocnrlkvl6dixeprgtv0xlrmwujhdy5qi6md71yupr4hwrgyj9ym1ev A6Aniket AmanNo ratings yet
- 1041 FullDocument6 pages1041 FullAdriano ItoNo ratings yet
- 11.ORoak Et Al 2012Document77 pages11.ORoak Et Al 2012MiaNo ratings yet
- 2016-Sung-Jin Park-Phototactic guidance of a tissue-engineered soft-robotic rayDocument6 pages2016-Sung-Jin Park-Phototactic guidance of a tissue-engineered soft-robotic rayitm.ggwpNo ratings yet
- Role of Genotype in The Cycle of Violence in Maltreated ChildrenDocument5 pagesRole of Genotype in The Cycle of Violence in Maltreated ChildrenMiquel AlaberniaNo ratings yet
- A General Framework For Analyzing Sustainability of Social-Ecological SystemsDocument5 pagesA General Framework For Analyzing Sustainability of Social-Ecological SystemsgranaditoshshNo ratings yet
- A General Framework For Analyzing Sustainability of Social-Ecological SystemsDocument4 pagesA General Framework For Analyzing Sustainability of Social-Ecological SystemsAlessandro Roberto de OliveiraNo ratings yet
- Iceland Fertility CousinsDocument5 pagesIceland Fertility CousinshalfdeadNo ratings yet
- Nature NeuropsicologiaDocument3 pagesNature NeuropsicologiaGabrielVásquezNo ratings yet
- Influence of Life Stress OnDocument4 pagesInfluence of Life Stress Ondeiab842728No ratings yet
- He Cken Berger 92008Document6 pagesHe Cken Berger 92008llerenasNo ratings yet
- Barriopedro 11Document6 pagesBarriopedro 11JxPNo ratings yet
- The Following Resources Related To This Article Are Available Online atDocument6 pagesThe Following Resources Related To This Article Are Available Online atChia-Lin Charles HoNo ratings yet
- Science 1201080Document5 pagesScience 1201080NeerajNo ratings yet
- Reports: Experimental Realization of Wheeler's Delayed-Choice Gedanken ExperimentDocument4 pagesReports: Experimental Realization of Wheeler's Delayed-Choice Gedanken ExperimentreflectiondarkNo ratings yet
- D C EliasScience 323, 610 (2009)Document5 pagesD C EliasScience 323, 610 (2009)Tania ChatterjeeNo ratings yet
- 2009 Science Jimenez CompleteDocument26 pages2009 Science Jimenez CompleteDarko PopovskiNo ratings yet
- Spatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesDocument6 pagesSpatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesJaqueline Godinez CamachoNo ratings yet
- 60.full Photochemical ProcessDocument5 pages60.full Photochemical ProcessDhanashri Rathod 18357No ratings yet
- High-Throughput Synthesis of Zeolitic Imidazolate Frameworks and Application To CO2 Capture (DRX)Document6 pagesHigh-Throughput Synthesis of Zeolitic Imidazolate Frameworks and Application To CO2 Capture (DRX)Annaíres AlmeidaNo ratings yet
- Catalytic: Cracking Acetic Acid To AceticDocument6 pagesCatalytic: Cracking Acetic Acid To AceticPawan NagarNo ratings yet
- Banerjee 2008Document6 pagesBanerjee 2008paulaNo ratings yet
- 2002 Irmof-2 EdusolDocument5 pages2002 Irmof-2 EdusolAlejandra AwimbaweNo ratings yet
- Cuprate Spin-Charge Order Observed in Quasi-Particle SpectrumDocument5 pagesCuprate Spin-Charge Order Observed in Quasi-Particle SpectrumAlejandra AwimbaweNo ratings yet
- A Modified Double Loop EPR Technique For Detection of Sensitization in Cast Stainless SteelDocument3 pagesA Modified Double Loop EPR Technique For Detection of Sensitization in Cast Stainless Steelcuentas ricardoNo ratings yet
- The Earth's Middle AtmosphereFrom EverandThe Earth's Middle AtmosphereW. L. GroseNo ratings yet
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocument5 pagesAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetNo ratings yet
- Abbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFDocument2 pagesAbbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFflashjetNo ratings yet
- Abbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFDocument5 pagesAbbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFflashjetNo ratings yet
- Nampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFDocument4 pagesNampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFflashjetNo ratings yet
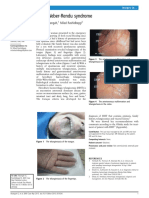
- Abangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeDocument2 pagesAbangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeflashjetNo ratings yet
- Nascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestDocument4 pagesNascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestflashjetNo ratings yet
- Nana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFDocument2 pagesNana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFflashjetNo ratings yet
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocument4 pagesAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetNo ratings yet
- Nascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntDocument3 pagesNascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntflashjetNo ratings yet
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocument5 pagesAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetNo ratings yet
- Namburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFDocument4 pagesNamburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFflashjetNo ratings yet
- Nathwani - 2013 - A Complicated Case of AnticoagulationDocument2 pagesNathwani - 2013 - A Complicated Case of AnticoagulationflashjetNo ratings yet
- Nascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesDocument3 pagesNascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesflashjetNo ratings yet
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocument4 pagesAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetNo ratings yet
- Narula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFDocument3 pagesNarula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFflashjetNo ratings yet
- Nasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeDocument3 pagesNasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeflashjetNo ratings yet
- Nascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseDocument5 pagesNascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseflashjetNo ratings yet
- Stroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437Document29 pagesStroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437MaryamNo ratings yet
- Nasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildDocument3 pagesNasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildflashjetNo ratings yet
- Nassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctDocument3 pagesNassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctflashjetNo ratings yet
- Idiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewDocument7 pagesIdiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewflashjetNo ratings yet
- Naveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarDocument4 pagesNaveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarflashjetNo ratings yet
- Natrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaDocument6 pagesNatrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaflashjetNo ratings yet
- Nassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirDocument3 pagesNassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirflashjetNo ratings yet
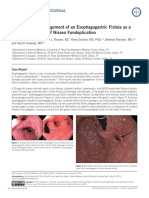
- Naveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationDocument2 pagesNaveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationflashjetNo ratings yet
- Nguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2Document36 pagesNguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2flashjetNo ratings yet
- Notes 200430 195226 0c2 PDFDocument19 pagesNotes 200430 195226 0c2 PDFflashjetNo ratings yet
- Notes 190915 213814 Ea7 PDFDocument1 pageNotes 190915 213814 Ea7 PDFflashjetNo ratings yet
- Naveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisDocument4 pagesNaveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisflashjetNo ratings yet
- Nguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisDocument4 pagesNguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisflashjetNo ratings yet
- Qualitative PPT 291118Document15 pagesQualitative PPT 291118Ixora MyNo ratings yet
- Essencial Natural Science Unit 1 TestDocument2 pagesEssencial Natural Science Unit 1 TestAlvaro Rizzo GonzálezNo ratings yet
- Sample exam questions on DNA, transcription, and translationDocument4 pagesSample exam questions on DNA, transcription, and translationLorraine M. Del RosarioNo ratings yet
- 1GS Cell PPT 2018Document73 pages1GS Cell PPT 2018pixiedustNo ratings yet
- The Cell Membrane: AP BiologyDocument53 pagesThe Cell Membrane: AP BiologythalesNo ratings yet
- Ashotinthearmora Hard Pill To Swallow?: Comparing Vaccine FormulationsDocument3 pagesAshotinthearmora Hard Pill To Swallow?: Comparing Vaccine FormulationsKaren SinorNo ratings yet
- Slip Test-5Document3 pagesSlip Test-5G.N. BhuvaneshwaranNo ratings yet
- Current Protocols in Molecular BiologyDocument11 pagesCurrent Protocols in Molecular BiologyMiriam Illescas0% (3)
- Fall 2021: Seminar 2: Cell Cycle, Mitosis and MeiosisDocument24 pagesFall 2021: Seminar 2: Cell Cycle, Mitosis and MeiosisChanceNo ratings yet
- Cell Structure and Function Test WorksheetDocument2 pagesCell Structure and Function Test WorksheetJohn Van Dave TaturoNo ratings yet
- Cell Cycle, mitosis worksheetDocument3 pagesCell Cycle, mitosis worksheetrosellevalerio2014No ratings yet
- Genomics Course: Basics of Genome SequencingDocument10 pagesGenomics Course: Basics of Genome Sequencingk.sachinNo ratings yet
- Types of MutationsDocument64 pagesTypes of MutationsI'm Cracked100% (1)
- MS-MS Analysis Programs - 2012 SlidesDocument14 pagesMS-MS Analysis Programs - 2012 SlidesJovanderson JacksonNo ratings yet
- Overview of Biochemical EndocrinologyDocument20 pagesOverview of Biochemical EndocrinologyGauri KashyapNo ratings yet
- Biocatalysis Questions and AnswersDocument9 pagesBiocatalysis Questions and Answerskumara guruparanNo ratings yet
- Gene transfer using retro and adeno virusesDocument17 pagesGene transfer using retro and adeno virusesVyshali PingleNo ratings yet
- 2020 GKS-G Available Universities & Fields of Study (English)Document348 pages2020 GKS-G Available Universities & Fields of Study (English)Rahma FadhilaNo ratings yet
- NURSING ANAPHY Lec Session #12 - SAS (NEW FORMAT) PDFDocument2 pagesNURSING ANAPHY Lec Session #12 - SAS (NEW FORMAT) PDFKriztel Andrei Lapac NavajaNo ratings yet
- 1. Chuyển hóa carbohydrate (metabolism of carbohydrates) : GlycolysisDocument20 pages1. Chuyển hóa carbohydrate (metabolism of carbohydrates) : GlycolysisAnh Tuyet NguyenNo ratings yet
- Drug DesignDocument38 pagesDrug DesignPhArMaCyGrAdUaTeSNo ratings yet
- Ethical Aspects of BiotechnologyDocument47 pagesEthical Aspects of Biotechnologyalfi alfathana100% (1)
- 473-Article Text-903-1-10-20181112Document122 pages473-Article Text-903-1-10-20181112unaiza100% (1)
- B.N.Bandodkar College Post-Reaccreditation ReportDocument47 pagesB.N.Bandodkar College Post-Reaccreditation Reportsdf6619No ratings yet
- The Chaleging BiofilmDocument58 pagesThe Chaleging BiofilmZayra Coral Ortiz OjedaNo ratings yet
- Genetic Markers: DNA Markers Provide High ResolutionDocument19 pagesGenetic Markers: DNA Markers Provide High ResolutionhotmarulitualinggaNo ratings yet