You might also like
- The Earth's Ionosphere: Plasma Physics and ElectrodynamicsFrom EverandThe Earth's Ionosphere: Plasma Physics and ElectrodynamicsNo ratings yet
- Singer Et Al2004Document6 pagesSinger Et Al2004Firuza RahimovaNo ratings yet
- Excitation of Orbital Angular Momentum Resonances in Helically Twisted Photonic Crystal FiberDocument4 pagesExcitation of Orbital Angular Momentum Resonances in Helically Twisted Photonic Crystal Fiberthillai saravananNo ratings yet
- Suppression of Rain and Snow by Urban and Industrial Air PollutionDocument6 pagesSuppression of Rain and Snow by Urban and Industrial Air PollutionDavidNo ratings yet
- Science (New York NY) 2003 TemelesDocument4 pagesScience (New York NY) 2003 TemelesmariaNo ratings yet
- Science 1201080Document5 pagesScience 1201080NeerajNo ratings yet
- Articulo Cientico 2Document5 pagesArticulo Cientico 2Elvis AndíaNo ratings yet
- 2002 Irmof-2 EdusolDocument5 pages2002 Irmof-2 EdusolAlejandra AwimbaweNo ratings yet
- Cuprate Spin-Charge Order Observed in Quasi-Particle SpectrumDocument5 pagesCuprate Spin-Charge Order Observed in Quasi-Particle SpectrumAlejandra AwimbaweNo ratings yet
- Suppression of Rain and Snow by Urban and Industrial Air PollutionDocument4 pagesSuppression of Rain and Snow by Urban and Industrial Air PollutionPedroNo ratings yet
- A Physical Map of 30,000 Human Genes: J. Am. Chem. Soc. 117, 4193 (1995) ), As DemonstratDocument3 pagesA Physical Map of 30,000 Human Genes: J. Am. Chem. Soc. 117, 4193 (1995) ), As DemonstratRahmat Eko SanjayaNo ratings yet
- Active Normal Faulting in The Upper Rhine Graben and Paleoseismic Identification of The 1356 Basel Earthquake.Document5 pagesActive Normal Faulting in The Upper Rhine Graben and Paleoseismic Identification of The 1356 Basel Earthquake.marceloahumada1983No ratings yet
- Simulating Dynamical Features of Escape Panic: Dirk Helbing, Illeâs Farkas & Tamaâs VicsekDocument4 pagesSimulating Dynamical Features of Escape Panic: Dirk Helbing, Illeâs Farkas & Tamaâs VicsektariqNo ratings yet
- A Logic-Gated Nanorobot For Targeted Transport of Molecular PayloadsDocument5 pagesA Logic-Gated Nanorobot For Targeted Transport of Molecular PayloadsJohnathonNo ratings yet
- Nuclei Get TAN LinesDocument3 pagesNuclei Get TAN LinesArpan NandyNo ratings yet
- 4D Electron Tomography ScienceDocument6 pages4D Electron Tomography Sciencehappynewyear11No ratings yet
- Ultrahigh Porosity in Metal-Organic Frameworks: 'KeeffeDocument6 pagesUltrahigh Porosity in Metal-Organic Frameworks: 'Keeffeクマール ヴァンツNo ratings yet
- Photonic Band Gap Guidance in Optical Fibers: J. C. Knight, J. Broeng, T. A. Birks, P. St. J. RussellDocument3 pagesPhotonic Band Gap Guidance in Optical Fibers: J. C. Knight, J. Broeng, T. A. Birks, P. St. J. RussellArnab BhowmikNo ratings yet
- High-Accuracy Ab Initio Rotation-Vibration Transitions For WaterDocument4 pagesHigh-Accuracy Ab Initio Rotation-Vibration Transitions For WaternchpascualNo ratings yet
- Superluminal and Slow Light Propagation in a Room-Temperature Solid - Science, 2003-200-2, BigelowDocument4 pagesSuperluminal and Slow Light Propagation in a Room-Temperature Solid - Science, 2003-200-2, BigelowRado JaNo ratings yet
- Chalfie 1994Document4 pagesChalfie 1994Basab BijayeeNo ratings yet
- The Following Resources Related To This Article Are Available Online atDocument5 pagesThe Following Resources Related To This Article Are Available Online atKoussa Monchi DalmarNo ratings yet
- Reports: Experimental Realization of Wheeler's Delayed-Choice Gedanken ExperimentDocument4 pagesReports: Experimental Realization of Wheeler's Delayed-Choice Gedanken ExperimentreflectiondarkNo ratings yet
- Science 2011 Vigny 1417 21 PDFDocument6 pagesScience 2011 Vigny 1417 21 PDFQuisiera Morir CantandoNo ratings yet
- 28 Sci 2007Document4 pages28 Sci 2007S GrixNo ratings yet
- Paper I PDFDocument6 pagesPaper I PDFErick FariasNo ratings yet
- Modelling The Evolution of Human Trail Systems: Letters To NatureDocument4 pagesModelling The Evolution of Human Trail Systems: Letters To NatureenmaNo ratings yet
- Boletin de Arquelologia PUCP No. 16 (2014) Número 16. Los Rostros de Huari Perspectivas Interregionales Sobre El Horizonte MedioDocument5 pagesBoletin de Arquelologia PUCP No. 16 (2014) Número 16. Los Rostros de Huari Perspectivas Interregionales Sobre El Horizonte MedioPareja Anyosa DanteNo ratings yet
- Optik 26Document13 pagesOptik 26z.umul9031No ratings yet
- The Following Resources Related To This Article Are Available Online atDocument5 pagesThe Following Resources Related To This Article Are Available Online atihzaoloanNo ratings yet
- + 2004 Atomic Transient RecorderDocument5 pages+ 2004 Atomic Transient RecorderDoktor transmisionesNo ratings yet
- Magnetic Skyrmions: Advances in Physics and Potential ApplicationsDocument5 pagesMagnetic Skyrmions: Advances in Physics and Potential ApplicationsAgtc TandayNo ratings yet
- Lieberman & Eisenberger (2009) Science PDFDocument2 pagesLieberman & Eisenberger (2009) Science PDFJulie ReNo ratings yet
- Self-Assembly at All Scales (Whitesides and Grzybowzki) - 1 PDFDocument5 pagesSelf-Assembly at All Scales (Whitesides and Grzybowzki) - 1 PDFAndrea Marie ComeoNo ratings yet
- Science 1213229Document5 pagesScience 12132291592162022No ratings yet
- Reducing The Dimensionality of Data With Neural Networks: ReportsDocument4 pagesReducing The Dimensionality of Data With Neural Networks: Reportsapi-25914596No ratings yet
- Science 1202393Document8 pagesScience 1202393CARLOS FRANCISCO MANTILLA MONTALVONo ratings yet
- Reducing The Dimensionality of Data With Neural Networks: ReportsDocument5 pagesReducing The Dimensionality of Data With Neural Networks: ReportsBruno Fabio Bedón VásquezNo ratings yet
- Improved Oxygen Reduction Activity On Pt3Ni Via Increased Surface Site AvailabilityDocument6 pagesImproved Oxygen Reduction Activity On Pt3Ni Via Increased Surface Site AvailabilityARIADNANo ratings yet
- Iceland Fertility CousinsDocument5 pagesIceland Fertility CousinshalfdeadNo ratings yet
- Visualizing Individual Nitrogen Dopants in Monolayer GrapheneDocument6 pagesVisualizing Individual Nitrogen Dopants in Monolayer Graphene김영찬No ratings yet
- HAARPDocument22 pagesHAARPOmar Valdivia MoralesNo ratings yet
- Science 2005 Nguyen 844 7Document5 pagesScience 2005 Nguyen 844 7Florin AndreiNo ratings yet
- 11.ORoak Et Al 2012Document77 pages11.ORoak Et Al 2012MiaNo ratings yet
- 2009 Science Jimenez CompleteDocument26 pages2009 Science Jimenez CompleteDarko PopovskiNo ratings yet
- Benazet El Al. Science (2009) A Self-Regulatory System of Interlinked SignalingDocument5 pagesBenazet El Al. Science (2009) A Self-Regulatory System of Interlinked SignalingnovrodNo ratings yet
- This Content Downloaded From 121.52.159.133 On Wed, 02 Jun 2021 09:58:24 UTCDocument8 pagesThis Content Downloaded From 121.52.159.133 On Wed, 02 Jun 2021 09:58:24 UTCShar KhanNo ratings yet
- Selyem - Holographically Controlled Three-Dimensional Atomic Population PatternsDocument10 pagesSelyem - Holographically Controlled Three-Dimensional Atomic Population PatternsJos JedanNo ratings yet
- Science 2005 Falke TalDocument21 pagesScience 2005 Falke TalBlueScholarNo ratings yet
- Buckler2009 PDFDocument6 pagesBuckler2009 PDFCarlitos MaldonadoNo ratings yet
- E.Behnke Et Al., Science, 319, (2008) - Spin Dependent WIMP Limits From A Bubble ChamberDocument4 pagesE.Behnke Et Al., Science, 319, (2008) - Spin Dependent WIMP Limits From A Bubble ChamberIvan FelisNo ratings yet
- Jobaria PDFDocument7 pagesJobaria PDFkallenhard1No ratings yet
- Science Aaa9633 PDFDocument4 pagesScience Aaa9633 PDFZoltan NadasdyNo ratings yet
- Self-Assembly Reading PDFDocument5 pagesSelf-Assembly Reading PDFYagmur İrem TemizlerNo ratings yet
- PhDThesisH Bartolf-DefenseNov2009Document24 pagesPhDThesisH Bartolf-DefenseNov2009Michelle CedNo ratings yet
- 2007 Ultrastrong and Stiff Layered Polymer Nanocomposites ScienceDocument5 pages2007 Ultrastrong and Stiff Layered Polymer Nanocomposites ScienceНаталья ГайдукNo ratings yet
- Bio Selective Membrane ElectrodeDocument6 pagesBio Selective Membrane Electrodeahmadalijee70No ratings yet
- Genomic RemodellingDocument2 pagesGenomic RemodellingDenys Quispe AlcocerNo ratings yet
- Electromagnetism ReferencesDocument7 pagesElectromagnetism ReferencesNivethithaa DhanrajNo ratings yet
- H.K. Moffatt - Magnetic Field Generation in Electrically Conducting Fluids: ReferencesDocument12 pagesH.K. Moffatt - Magnetic Field Generation in Electrically Conducting Fluids: ReferencesOsdfmNo ratings yet
- Sec Bhavdata FullDocument71 pagesSec Bhavdata FullarchsarkNo ratings yet
- Journal of Statistical Software: Learning Bayesian Networks With The Bnlearn R PackageDocument22 pagesJournal of Statistical Software: Learning Bayesian Networks With The Bnlearn R PackagearchsarkNo ratings yet
- Journal of Statistical Software: Learning Bayesian Networks With The Bnlearn R PackageDocument22 pagesJournal of Statistical Software: Learning Bayesian Networks With The Bnlearn R PackagearchsarkNo ratings yet
- Science 2005Document43 pagesScience 2005archsarkNo ratings yet
- Journal of Statistical Software: Learning Bayesian Networks With The Bnlearn R PackageDocument22 pagesJournal of Statistical Software: Learning Bayesian Networks With The Bnlearn R PackagearchsarkNo ratings yet
- Reconstructing signaling networks from single-cell dataDocument8 pagesReconstructing signaling networks from single-cell dataarchsarkNo ratings yet
- Reconstructing signaling networks from single-cell dataDocument8 pagesReconstructing signaling networks from single-cell dataarchsarkNo ratings yet
- Finance As An Industrial ScienceDocument2 pagesFinance As An Industrial SciencearchsarkNo ratings yet
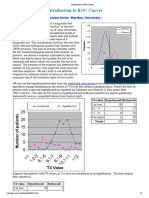
- Introduction To ROC Curves PDFDocument2 pagesIntroduction To ROC Curves PDFarchsarkNo ratings yet
- Computer Science and Applications - English OnlyDocument8 pagesComputer Science and Applications - English OnlyUma MaheshNo ratings yet
- Temporal EvolutionDocument18 pagesTemporal EvolutionarchsarkNo ratings yet
- Minkowski DistanceDocument2 pagesMinkowski Distancearchsark100% (1)
- Algo Trading - Quant Congress USA 2011Document58 pagesAlgo Trading - Quant Congress USA 2011Sukju HwangNo ratings yet
- Agriculture 03 00516Document20 pagesAgriculture 03 00516archsarkNo ratings yet
- University Grants Commission - NETDocument1 pageUniversity Grants Commission - NETarchsarkNo ratings yet
- Mathematical Operators: Range: 2200-22FFDocument8 pagesMathematical Operators: Range: 2200-22FFarchsarkNo ratings yet
- SOCIOLOGY G93.3306-001 Event History AnalysisDocument15 pagesSOCIOLOGY G93.3306-001 Event History AnalysisarchsarkNo ratings yet
- How-To - Install CDH On Mac OSX 10Document20 pagesHow-To - Install CDH On Mac OSX 10archsarkNo ratings yet
- NET Syllabus Computer Science ITDocument11 pagesNET Syllabus Computer Science ITnetmat100% (4)
- NET EXAM Paper 1Document27 pagesNET EXAM Paper 1shahid ahmed laskar100% (4)
- D 8706 Paper IiiDocument40 pagesD 8706 Paper IiiashashekarNo ratings yet
- Building A Smart Engineering Team... Differently!Document3 pagesBuilding A Smart Engineering Team... Differently!archsarkNo ratings yet
- Chapter 17 Projects: 1. (After 17.3) Changing Color and AlignmentDocument14 pagesChapter 17 Projects: 1. (After 17.3) Changing Color and AlignmentarchsarkNo ratings yet
- Efron MorrisDocument10 pagesEfron MorrisarchsarkNo ratings yet
- Legend of Nueva VizcayaDocument10 pagesLegend of Nueva VizcayaAndreanna Maria100% (1)
- B Ed 3 Sem Physics Teaching Group C Science 1 Paper 1 Summer 2018Document3 pagesB Ed 3 Sem Physics Teaching Group C Science 1 Paper 1 Summer 2018Rrr KkkNo ratings yet
- Managed Pressure Drilling MPD BrochureDocument5 pagesManaged Pressure Drilling MPD Brochureswaala4realNo ratings yet
- Welcome Students!: Week 8 (3 Quarter)Document28 pagesWelcome Students!: Week 8 (3 Quarter)Erika Lloren Luyun-GaliaNo ratings yet
- Differences in Left Ventricular and Left Atrial FuDocument10 pagesDifferences in Left Ventricular and Left Atrial FueugeniaNo ratings yet
- Thalassemia: Submitted By: Jovan Pierre C. Ouano Submitted To: Mark Gil T. DacutanDocument8 pagesThalassemia: Submitted By: Jovan Pierre C. Ouano Submitted To: Mark Gil T. DacutanJvnpierre AberricanNo ratings yet
- Terpin Hydrate Oral SolutionDocument1 pageTerpin Hydrate Oral SolutionAbelard Maria EscrivaNo ratings yet
- NTPC Training ReportDocument83 pagesNTPC Training ReportAbhishek Mittal100% (2)
- Auomotivr Fuel TanksDocument9 pagesAuomotivr Fuel Tankscolumba_mcNo ratings yet
- Mass DisasterDocument70 pagesMass DisasterJoseph RadovanNo ratings yet
- Understanding The Nature of ReligionDocument41 pagesUnderstanding The Nature of Religioneden l. abadNo ratings yet
- Hook and Eye-Stories Form The MarginsDocument220 pagesHook and Eye-Stories Form The MarginsJefferson LiinNo ratings yet
- 2a CTRL DecDocument75 pages2a CTRL Decramanathan balamoorthyNo ratings yet
- Fit For WashingDocument85 pagesFit For WashingIyaka YoeNo ratings yet
- FCV585 FCV620 Operator's Manual G 1-7-11Document57 pagesFCV585 FCV620 Operator's Manual G 1-7-11Katia QuispeNo ratings yet
- 30-300 MHZ Transmission Line Trainer Tla05Document1 page30-300 MHZ Transmission Line Trainer Tla05Kishor HandoreNo ratings yet
- Erotic MassageDocument113 pagesErotic MassageMahmoud El Mohamdy75% (4)
- Week 7: Nurses Role in Disaster: Home Mitigation and PreparednessDocument10 pagesWeek 7: Nurses Role in Disaster: Home Mitigation and PreparednessRose Ann LacuarinNo ratings yet
- Pantheon-Katalog Eng WebDocument16 pagesPantheon-Katalog Eng WebJoe DoeNo ratings yet
- PSUTIL Documentation for Monitoring System ResourcesDocument77 pagesPSUTIL Documentation for Monitoring System ResourcesImam RiadiNo ratings yet
- VSD Power Supply Connections and EarthingDocument4 pagesVSD Power Supply Connections and EarthingHumaid ShaikhNo ratings yet
- Conduction Calorimetric Investigation of The Effect of Retarders On The Hydration of Portland CementDocument15 pagesConduction Calorimetric Investigation of The Effect of Retarders On The Hydration of Portland CementAlfredo Landaverde GarcíaNo ratings yet
- Well Rounded.: 360 CassetteDocument12 pagesWell Rounded.: 360 CassetteMonty Va Al MarNo ratings yet
- EniSpA - The Corporate Strategy of An International Energy Major PDFDocument24 pagesEniSpA - The Corporate Strategy of An International Energy Major PDFAnonymous 9ZMbuR75% (4)
- Lecture No.3 Part 1 (Fan)Document6 pagesLecture No.3 Part 1 (Fan)Mohsen HassanNo ratings yet
- 5020-Article Text-10917-1-10-20220808Document9 pages5020-Article Text-10917-1-10-20220808indah rumah4No ratings yet
- Bentley Bentayga BrochureDocument9 pagesBentley Bentayga BrochureGerry CalàNo ratings yet
- Petford & Atherton., 1996Document31 pagesPetford & Atherton., 1996Ivan Hagler Becerra VasquezNo ratings yet
- SGDJ PDFDocument334 pagesSGDJ PDFDouglas Rogério de CastroNo ratings yet
- Textile Internship Report AlokDocument39 pagesTextile Internship Report AlokRahul TelangNo ratings yet
- Microsoft Azure Infrastructure Services for Architects: Designing Cloud SolutionsFrom EverandMicrosoft Azure Infrastructure Services for Architects: Designing Cloud SolutionsNo ratings yet
- AWS Certified Cloud Practitioner Study Guide: CLF-C01 ExamFrom EverandAWS Certified Cloud Practitioner Study Guide: CLF-C01 ExamRating: 5 out of 5 stars5/5 (1)
- Computer Systems and Networking Guide: A Complete Guide to the Basic Concepts in Computer Systems, Networking, IP Subnetting and Network SecurityFrom EverandComputer Systems and Networking Guide: A Complete Guide to the Basic Concepts in Computer Systems, Networking, IP Subnetting and Network SecurityRating: 4.5 out of 5 stars4.5/5 (13)
- CCNA: 3 in 1- Beginner's Guide+ Tips on Taking the Exam+ Simple and Effective Strategies to Learn About CCNA (Cisco Certified Network Associate) Routing And Switching CertificationFrom EverandCCNA: 3 in 1- Beginner's Guide+ Tips on Taking the Exam+ Simple and Effective Strategies to Learn About CCNA (Cisco Certified Network Associate) Routing And Switching CertificationNo ratings yet
- ITIL 4: Digital and IT strategy: Reference and study guideFrom EverandITIL 4: Digital and IT strategy: Reference and study guideRating: 5 out of 5 stars5/5 (1)
- The Compete Ccna 200-301 Study Guide: Network Engineering EditionFrom EverandThe Compete Ccna 200-301 Study Guide: Network Engineering EditionRating: 5 out of 5 stars5/5 (4)
- Amazon Web Services (AWS) Interview Questions and AnswersFrom EverandAmazon Web Services (AWS) Interview Questions and AnswersRating: 4.5 out of 5 stars4.5/5 (3)
- The CompTIA Network+ Computing Technology Industry Association Certification N10-008 Study Guide: Hi-Tech Edition: Proven Methods to Pass the Exam with Confidence - Practice Test with AnswersFrom EverandThe CompTIA Network+ Computing Technology Industry Association Certification N10-008 Study Guide: Hi-Tech Edition: Proven Methods to Pass the Exam with Confidence - Practice Test with AnswersNo ratings yet
- The Ultimate Kali Linux Book - Second Edition: Perform advanced penetration testing using Nmap, Metasploit, Aircrack-ng, and EmpireFrom EverandThe Ultimate Kali Linux Book - Second Edition: Perform advanced penetration testing using Nmap, Metasploit, Aircrack-ng, and EmpireNo ratings yet
- Computer Networking: The Complete Beginner's Guide to Learning the Basics of Network Security, Computer Architecture, Wireless Technology and Communications Systems (Including Cisco, CCENT, and CCNA)From EverandComputer Networking: The Complete Beginner's Guide to Learning the Basics of Network Security, Computer Architecture, Wireless Technology and Communications Systems (Including Cisco, CCENT, and CCNA)Rating: 4 out of 5 stars4/5 (4)
- Evaluation of Some Websites that Offer Virtual Phone Numbers for SMS Reception and Websites to Obtain Virtual Debit/Credit Cards for Online Accounts VerificationsFrom EverandEvaluation of Some Websites that Offer Virtual Phone Numbers for SMS Reception and Websites to Obtain Virtual Debit/Credit Cards for Online Accounts VerificationsRating: 5 out of 5 stars5/5 (1)
- Computer Networking: The Complete Guide to Understanding Wireless Technology, Network Security, Computer Architecture and Communications Systems (Including Cisco, CCNA and CCENT)From EverandComputer Networking: The Complete Guide to Understanding Wireless Technology, Network Security, Computer Architecture and Communications Systems (Including Cisco, CCNA and CCENT)No ratings yet
- Designing and Building Security Operations CenterFrom EverandDesigning and Building Security Operations CenterRating: 3 out of 5 stars3/5 (3)
- ITIL® 4 Direct, Plan and Improve (DPI): Your companion to the ITIL 4 Managing Professional and Strategic Leader DPI certificationFrom EverandITIL® 4 Direct, Plan and Improve (DPI): Your companion to the ITIL 4 Managing Professional and Strategic Leader DPI certificationNo ratings yet
- Introduction to Cyber-Warfare: A Multidisciplinary ApproachFrom EverandIntroduction to Cyber-Warfare: A Multidisciplinary ApproachRating: 4.5 out of 5 stars4.5/5 (3)
- AWS Certified Solutions Architect Study Guide: Associate SAA-C02 ExamFrom EverandAWS Certified Solutions Architect Study Guide: Associate SAA-C02 ExamNo ratings yet
- ITIL® 4 Create, Deliver and Support (CDS): Your companion to the ITIL 4 Managing Professional CDS certificationFrom EverandITIL® 4 Create, Deliver and Support (CDS): Your companion to the ITIL 4 Managing Professional CDS certificationRating: 5 out of 5 stars5/5 (2)
- Advanced Antenna Systems for 5G Network Deployments: Bridging the Gap Between Theory and PracticeFrom EverandAdvanced Antenna Systems for 5G Network Deployments: Bridging the Gap Between Theory and PracticeRating: 5 out of 5 stars5/5 (1)