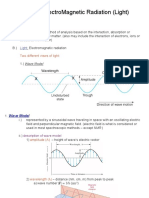
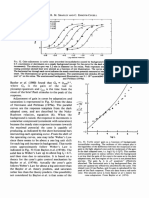
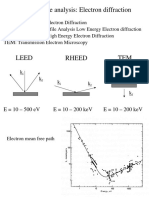
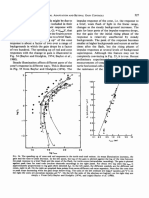
You might also like
- SCL 90R Scales AnalysisDocument9 pagesSCL 90R Scales AnalysisCarolina Almonacid60% (10)
- The Beginners Bible Coloring Book by JPR504Document37 pagesThe Beginners Bible Coloring Book by JPR504Ernesto Carmona Parra100% (1)
- More Notes of A Dirty Old Man (Converted FromDocument499 pagesMore Notes of A Dirty Old Man (Converted FromŽeljko ĐurićNo ratings yet
- ISnack - Sec A Group6Document11 pagesISnack - Sec A Group6Avik BorahNo ratings yet
- Vibrational Behaviour of The Turbo Generator Stator End Winding in CaseDocument12 pagesVibrational Behaviour of The Turbo Generator Stator End Winding in Casekoohestani_afshin50% (2)
- Chapter 6 Coming of SpainDocument4 pagesChapter 6 Coming of SpainJayvee MacapagalNo ratings yet
- Physics 571 Lecture #23: 1 Acousto-Optic Modulators (AOM)Document4 pagesPhysics 571 Lecture #23: 1 Acousto-Optic Modulators (AOM)ss626No ratings yet
- Int El Vib Rot: Chapter 10 - 1Document8 pagesInt El Vib Rot: Chapter 10 - 1rajasekaransj6455No ratings yet
- Measurement of The Lorenz-Mie Scattering of A Single Particle: Polystyrene LatexDocument4 pagesMeasurement of The Lorenz-Mie Scattering of A Single Particle: Polystyrene LatexNenn RmcNo ratings yet
- VISUAL ADAPTATION AND RETINAL GAIN CONTROLSDocument1 pageVISUAL ADAPTATION AND RETINAL GAIN CONTROLSAkicaNo ratings yet
- L03 MRI Part 2Document10 pagesL03 MRI Part 2lolpoll771No ratings yet
- Visual adaptation and retinal gain controlsDocument1 pageVisual adaptation and retinal gain controlsAkicaNo ratings yet
- Seismic Reflection Notes: Lithoprobe Slide SetDocument13 pagesSeismic Reflection Notes: Lithoprobe Slide SetstankomirNo ratings yet
- Do Not: 5. Gain Control in PhotoreceptorsDocument1 pageDo Not: 5. Gain Control in PhotoreceptorsAkicaNo ratings yet
- Locking of Hene Laser Modes Induced by Synchronous Intracavity ModulationDocument3 pagesLocking of Hene Laser Modes Induced by Synchronous Intracavity ModulationMohamad paidarNo ratings yet
- Generation of Squeezed States of Light: A Critical Discussion SDocument4 pagesGeneration of Squeezed States of Light: A Critical Discussion SJai singhNo ratings yet
- Interference in Thin FilmDocument56 pagesInterference in Thin FilmjameshrclassesNo ratings yet
- Mreznica - 74Document1 pageMreznica - 74AkicaNo ratings yet
- Module-I-Wave Optics With ProblmesDocument29 pagesModule-I-Wave Optics With Problmeshk01061903No ratings yet
- 421 821 Chapter 6Document17 pages421 821 Chapter 6April Mergelle LapuzNo ratings yet
- EG268EGA229EG238 Vibration Lab Sheet 2019Document6 pagesEG268EGA229EG238 Vibration Lab Sheet 2019classwizcasio12345No ratings yet
- Engineering Physics (PHY 1051)Document11 pagesEngineering Physics (PHY 1051)Harshita GauravNo ratings yet
- Oscillator Phase Noise: A Tutorial: Thomas H. Lee, Member, IEEE, and Ali Hajimiri, Member, IEEEDocument11 pagesOscillator Phase Noise: A Tutorial: Thomas H. Lee, Member, IEEE, and Ali Hajimiri, Member, IEEEvirendra_verma5No ratings yet
- Gouy Phase ShiftDocument4 pagesGouy Phase ShiftGörkem VarolNo ratings yet
- MQP Ans 01Document16 pagesMQP Ans 01singhkshama3404No ratings yet
- Schalaw Townes Condition (Lasers)Document3 pagesSchalaw Townes Condition (Lasers)ASHWINI100% (3)
- Material Characterization TechniquesDocument28 pagesMaterial Characterization TechniquesAmal ShajiNo ratings yet
- Tpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IDocument1 pageTpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IAkicaNo ratings yet
- Physics2 3 (Optics)Document21 pagesPhysics2 3 (Optics)zuuuNo ratings yet
- 2 Deconvolution PDFDocument112 pages2 Deconvolution PDFFelipe CorrêaNo ratings yet
- Lasers and Semiconductors: ContentDocument22 pagesLasers and Semiconductors: ContentAgus LeonardiNo ratings yet
- 6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oDocument1 page6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oAkicaNo ratings yet
- 06 Diffraction Basics IIDocument28 pages06 Diffraction Basics IIAli HuertaNo ratings yet
- Sunspot TRDocument4 pagesSunspot TRAnonymous b2k9ABe7eNo ratings yet
- Waves 3Document6 pagesWaves 3krichenkyandex.ruNo ratings yet
- Electron diffraction techniques for surface structure analysisDocument22 pagesElectron diffraction techniques for surface structure analysis陳帥睿No ratings yet
- MSP-705 Non Linear Optics - Notes 1Document95 pagesMSP-705 Non Linear Optics - Notes 1ArathyNo ratings yet
- Copying of Quantum Information by Means of A Quantum Amplifier (Laser Physics Letters, Vol. 3, Issue 2) (2006)Document4 pagesCopying of Quantum Information by Means of A Quantum Amplifier (Laser Physics Letters, Vol. 3, Issue 2) (2006)sepot24093No ratings yet
- Optical Fibre Types and Light PropagationDocument39 pagesOptical Fibre Types and Light Propagationteam 8No ratings yet
- Barrington LeighFeb01Document12 pagesBarrington LeighFeb01Natan SanglardNo ratings yet
- Pearson Edexcel IAL Physics Waves and Electricity Lecture NotesDocument18 pagesPearson Edexcel IAL Physics Waves and Electricity Lecture NotesBryan YeohNo ratings yet
- F33C06: Atoms, Photons and Elementary Particles: 1 RadioactivityDocument9 pagesF33C06: Atoms, Photons and Elementary Particles: 1 RadioactivityNina BrownNo ratings yet
- Vibrational SpectrosDocument42 pagesVibrational SpectrosAbhishekNo ratings yet
- On The Relation Between The Photoactivation Energy and The Absorbance Spectrum of Visual PigmentsDocument6 pagesOn The Relation Between The Photoactivation Energy and The Absorbance Spectrum of Visual PigmentsOctavio HernándezNo ratings yet
- 4.3. Horizontal Cells: F I I FDocument1 page4.3. Horizontal Cells: F I I FAkicaNo ratings yet
- Study of Various Designs of Solid State Laser Oscillator: Kireet Semwal, S. C. BhattDocument12 pagesStudy of Various Designs of Solid State Laser Oscillator: Kireet Semwal, S. C. BhattIOSRJEN : hard copy, certificates, Call for Papers 2013, publishing of journalNo ratings yet
- Rassejanie Mandelshtama BrilljuenaDocument55 pagesRassejanie Mandelshtama BrilljuenaNəzrin SultanNo ratings yet
- Optical Communication Full PDFDocument137 pagesOptical Communication Full PDFadityavishwakarma7434No ratings yet
- The Reflection Behaviours of Elliptical Cylinder-Loaded Monopole Antenna With Cylindrical BalunDocument4 pagesThe Reflection Behaviours of Elliptical Cylinder-Loaded Monopole Antenna With Cylindrical Balunfakher hamoucheNo ratings yet
- Radiation Loss 2Document6 pagesRadiation Loss 2torikulNo ratings yet
- Et Al., 1982) - This: M. Shapley and C. Enroth-CugellDocument1 pageEt Al., 1982) - This: M. Shapley and C. Enroth-CugellAkicaNo ratings yet
- Retina - 10Document1 pageRetina - 10AkicaNo ratings yet
- Am2c09063 Si 003Document13 pagesAm2c09063 Si 003Neurodynamics (NeuroDynamic Software)No ratings yet
- Antenna and Space Telecommunication HND 2020 SESSIONDocument5 pagesAntenna and Space Telecommunication HND 2020 SESSIONJeanFrancoisAtemengueEbanga100% (1)
- UNIT I - Wave OpticsDocument33 pagesUNIT I - Wave Opticsmirae kangNo ratings yet
- L04 MRI Part 3Document21 pagesL04 MRI Part 3lolpoll771No ratings yet
- 421 821 Chapter 6Document14 pages421 821 Chapter 6Marie Kris NogaNo ratings yet
- Sonar Introduction 2012 Compressed PDFDocument11 pagesSonar Introduction 2012 Compressed PDFBrenno Moura MouraNo ratings yet
- Flicker Rate TV Weapon Experiment On MonkeysDocument4 pagesFlicker Rate TV Weapon Experiment On Monkeysmarko2222No ratings yet
- Óptica de Fourier 2fDocument4 pagesÓptica de Fourier 2fSF EnriqueNo ratings yet
- Oxford Optics Notes 2nd YearDocument64 pagesOxford Optics Notes 2nd YearGörkem VarolNo ratings yet
- Seismic Sources GeophysicsDocument25 pagesSeismic Sources GeophysicsbadrhashmiNo ratings yet
- Amplitude Within The Fresnel Zone For The Zero-Offset CaseDocument11 pagesAmplitude Within The Fresnel Zone For The Zero-Offset CaseSaurav VijhNo ratings yet
- Fluorescence Microscopy With Diffraction Resolution Barrier Broken by Stimulated EmissionDocument5 pagesFluorescence Microscopy With Diffraction Resolution Barrier Broken by Stimulated EmissionAMNo ratings yet
- Mreznica - 80Document1 pageMreznica - 80AkicaNo ratings yet
- R. M. Shapley and C. Enroth-Cugell: Et AlDocument1 pageR. M. Shapley and C. Enroth-Cugell: Et AlAkicaNo ratings yet
- Mreznica - 73Document1 pageMreznica - 73AkicaNo ratings yet
- Mreznica - 79Document1 pageMreznica - 79AkicaNo ratings yet
- Mreznica - 78Document1 pageMreznica - 78AkicaNo ratings yet
- Visual Adaptation and Retinal Gain ControlsDocument1 pageVisual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- Mreznica - 74Document1 pageMreznica - 74AkicaNo ratings yet
- Visual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)Document1 pageVisual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)AkicaNo ratings yet
- Tpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IDocument1 pageTpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IAkicaNo ratings yet
- Et Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30Document1 pageEt Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30AkicaNo ratings yet
- Do Not: 5. Gain Control in PhotoreceptorsDocument1 pageDo Not: 5. Gain Control in PhotoreceptorsAkicaNo ratings yet
- Mreznica - 66Document1 pageMreznica - 66AkicaNo ratings yet
- Z,, (Co) Y.L (Co) I, (O ) I, (Co) : Visual Adaptation and Retinal Gain ControlsDocument1 pageZ,, (Co) Y.L (Co) I, (O ) I, (Co) : Visual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- Mreznica - 71Document1 pageMreznica - 71AkicaNo ratings yet
- Mreznica - 60Document1 pageMreznica - 60AkicaNo ratings yet
- 6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oDocument1 page6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oAkicaNo ratings yet
- Mreznica - 61Document1 pageMreznica - 61AkicaNo ratings yet
- Mreznica - 59Document1 pageMreznica - 59AkicaNo ratings yet
- Time Course of Gain Adjustment in Retinal Ganglion CellsDocument1 pageTime Course of Gain Adjustment in Retinal Ganglion CellsAkicaNo ratings yet
- 4.3. Horizontal Cells: F I I FDocument1 page4.3. Horizontal Cells: F I I FAkicaNo ratings yet
- Mreznica - 58Document1 pageMreznica - 58AkicaNo ratings yet
- Mreznica - 55Document1 pageMreznica - 55AkicaNo ratings yet
- Retina - 53 PDFDocument1 pageRetina - 53 PDFAkicaNo ratings yet
- Retina - 52Document1 pageRetina - 52AkicaNo ratings yet
- Retina - 52Document1 pageRetina - 52AkicaNo ratings yet
- Retina - 52Document1 pageRetina - 52AkicaNo ratings yet
- Visual Adaptation and Retinal Gain ControlsDocument1 pageVisual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- Retina - 51Document1 pageRetina - 51AkicaNo ratings yet
- 3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsDocument1 page3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsAkicaNo ratings yet
- Examen Tema 3 Ingles EnzoDocument3 pagesExamen Tema 3 Ingles EnzoMartinEnzo GuerraNo ratings yet
- High risk patient dental questionsDocument3 pagesHigh risk patient dental questionsFoysal SirazeeNo ratings yet
- The Madhya Pradesh Public Trusts Act 1951Document13 pagesThe Madhya Pradesh Public Trusts Act 1951Zenith ChhablaniNo ratings yet
- Module 1 MC4 LOGIC CRITICAL THINKINGDocument6 pagesModule 1 MC4 LOGIC CRITICAL THINKINGAubrey Ante100% (1)
- Target Mature Men with Luxury Watch MarketingDocument13 pagesTarget Mature Men with Luxury Watch MarketingAndriDwisondiNo ratings yet
- Talent ManagementDocument8 pagesTalent Managementyared haftuNo ratings yet
- The Future of The Past: The Contemporary Significance of The Nouvelle ThéologieDocument15 pagesThe Future of The Past: The Contemporary Significance of The Nouvelle ThéologiefesousacostaNo ratings yet
- 1988 Ojhri Camp Disaster in PakistanDocument3 pages1988 Ojhri Camp Disaster in PakistanranasohailiqbalNo ratings yet
- Leontine O'Gorman, RSCJDocument4 pagesLeontine O'Gorman, RSCJuptickdarter0rNo ratings yet
- Basic Accounting Lecture 05202018Document188 pagesBasic Accounting Lecture 05202018Rheea de los SantosNo ratings yet
- Duration of Labor: Group1Document19 pagesDuration of Labor: Group1ABEGAIL BALLORANNo ratings yet
- Finale 2005b - (Holly Mass of ST Peter DamianDocument6 pagesFinale 2005b - (Holly Mass of ST Peter DamianArmelNo ratings yet
- 01 FontanesiDocument7 pages01 FontanesizegorszyNo ratings yet
- TFN RVWDocument145 pagesTFN RVWmftaganasNo ratings yet
- SAT Writing Notes CollegeboardDocument43 pagesSAT Writing Notes CollegeboardEric JoNo ratings yet
- Soil MechanicsDocument117 pagesSoil MechanicsChiranjaya HulangamuwaNo ratings yet
- Clustered File Systems With de Series ProductsDocument28 pagesClustered File Systems With de Series ProductsRaulcb1No ratings yet
- American Folk Song Lesson PlanDocument1 pageAmerican Folk Song Lesson Planapi-2844060150% (1)
- Chapter5 EthDocument30 pagesChapter5 EthVictoriasecret PerfumeNo ratings yet
- SafeNet Ikey4000 PB (En) WebDocument2 pagesSafeNet Ikey4000 PB (En) WebbasharaccountNo ratings yet
- 05 AHP and Scoring ModelsDocument32 pages05 AHP and Scoring ModelsIhjaz VarikkodanNo ratings yet
- Sourcing Rules in PurchasingDocument21 pagesSourcing Rules in Purchasingpradhan.neeladriNo ratings yet
- Csir Ugc Net - Life Sciences - Free Sample TheoryDocument12 pagesCsir Ugc Net - Life Sciences - Free Sample TheorySyamala Natarajan100% (1)
- Book of AbstractsDocument156 pagesBook of Abstractsdragance106No ratings yet