You might also like
- 4.3. Horizontal Cells: F I I FDocument1 page4.3. Horizontal Cells: F I I FAkicaNo ratings yet
- Mreznica - 60Document1 pageMreznica - 60AkicaNo ratings yet
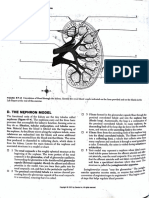
- D. The Ne Phron Model: Ul'lnary TructuresDocument6 pagesD. The Ne Phron Model: Ul'lnary TructuresCatherine BelonioNo ratings yet
- Ai. (1971) - The Cone Pigment Has Been Only BleachedDocument1 pageAi. (1971) - The Cone Pigment Has Been Only BleachedAkicaNo ratings yet
- Tpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IDocument1 pageTpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IAkicaNo ratings yet
- Mreznica - 61Document1 pageMreznica - 61AkicaNo ratings yet
- The Handbook of Infrared and Raman Characteristic Frequencies of Organic MoleculesFrom EverandThe Handbook of Infrared and Raman Characteristic Frequencies of Organic MoleculesRating: 5 out of 5 stars5/5 (2)
- TRDocument10 pagesTRMelanie MartinssonNo ratings yet
- LABManualDocument3 pagesLABManualChriszaine PerezNo ratings yet
- Exercise 15 Serial Transverse Section of A 24 Hour Chick Embryo PDFDocument5 pagesExercise 15 Serial Transverse Section of A 24 Hour Chick Embryo PDFMichaela FayeNo ratings yet
- Introduction to Feynman Diagrams: International Series of Monographs in Natural PhilosophyFrom EverandIntroduction to Feynman Diagrams: International Series of Monographs in Natural PhilosophyRating: 5 out of 5 stars5/5 (1)
- Neural Networks and Applications TutorialDocument45 pagesNeural Networks and Applications Tutorialvane-16No ratings yet
- Visual Adaptation and Retinal Gain ControlsDocument1 pageVisual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- Development of The Cerebral Cortex: Forming The Cortical StructureDocument2 pagesDevelopment of The Cerebral Cortex: Forming The Cortical StructureMouhamad Al-kurdiNo ratings yet
- Vaginal Smear Study Reveals Relationship Between Sexual Receptivity and Ovulation in RatsDocument9 pagesVaginal Smear Study Reveals Relationship Between Sexual Receptivity and Ovulation in RatsNatasha LamarkNo ratings yet
- Structure and Properties Oi Isotactic PolypropyleneDocument12 pagesStructure and Properties Oi Isotactic PolypropyleneNguyễn Mạnh TuấnNo ratings yet
- The Odor of Optical Isomers An Experiment in Organic ChemistryDocument2 pagesThe Odor of Optical Isomers An Experiment in Organic Chemistrygeovani2No ratings yet
- STPM Trial 2010 Bio 2 KelantanDocument8 pagesSTPM Trial 2010 Bio 2 Kelantanone kilometerNo ratings yet
- Adiabatic Motionqt0r64b33dDocument54 pagesAdiabatic Motionqt0r64b33deduardovb2023No ratings yet
- Week 1 BlankDocument3 pagesWeek 1 BlankAbraham WalkthewokNo ratings yet
- tmp7662 TMPDocument3 pagestmp7662 TMPFrontiersNo ratings yet
- Week 3: DNA ReplicationDocument39 pagesWeek 3: DNA ReplicationKEVIN SUNNo ratings yet
- Nuclear Structure and Heavy-Ion DynamicsFrom EverandNuclear Structure and Heavy-Ion DynamicsL. MorettoNo ratings yet
- Ab Initio Molecular Orbital Theory Warren HehreDocument8 pagesAb Initio Molecular Orbital Theory Warren HehrepedroNo ratings yet
- Artigo Do Corey Sobre Adição de LiAlH4 em Alcoóis PropargílicosDocument3 pagesArtigo Do Corey Sobre Adição de LiAlH4 em Alcoóis PropargílicosNatan FilippiNo ratings yet
- Blackshear 1956 NACA The Growth of Disturbances in A Flame Generated Shear RegionDocument11 pagesBlackshear 1956 NACA The Growth of Disturbances in A Flame Generated Shear RegionPrem ChandNo ratings yet
- "' Itry: Gan BacilliDocument13 pages"' Itry: Gan BacilliSandra PiedadeNo ratings yet
- Retina - 40Document1 pageRetina - 40AkicaNo ratings yet
- University of California Los Angeles .. - ,,-. - .: Approved For Release Cia-Rdp96-007B7R0004000Booic RDDocument6 pagesUniversity of California Los Angeles .. - ,,-. - .: Approved For Release Cia-Rdp96-007B7R0004000Booic RDChadNo ratings yet
- Band Structure and Transport Properties of Some 3-5 CompoundsDocument13 pagesBand Structure and Transport Properties of Some 3-5 CompoundsJose Leon GonzalezNo ratings yet
- Optical Fiber Rotation SensingFrom EverandOptical Fiber Rotation SensingWilliam K. BurnsRating: 3 out of 5 stars3/5 (1)
- An Essential Role For NF-KB in Preventing TNF-a-Induced Cell DeathDocument3 pagesAn Essential Role For NF-KB in Preventing TNF-a-Induced Cell DeathJuan manuel jiménez estradaNo ratings yet
- Chap 4 PROBLEMSDocument3 pagesChap 4 PROBLEMSsaubhagyamoharana62No ratings yet
- BROADHURST, M. G., 1962, An Analysis of The Solid Phase Behavior of The Normal ParaffinsDocument9 pagesBROADHURST, M. G., 1962, An Analysis of The Solid Phase Behavior of The Normal ParaffinsDiogo AlvesNo ratings yet
- Polymer Conformation and Configuration: A Polytechnic Press of the Polytechnic Institute of Brooklyn BookFrom EverandPolymer Conformation and Configuration: A Polytechnic Press of the Polytechnic Institute of Brooklyn BookNo ratings yet
- Structure of Molecules and Internal Rotation: Physical Chemistry: A Series of MonographsFrom EverandStructure of Molecules and Internal Rotation: Physical Chemistry: A Series of MonographsNo ratings yet
- (A) Parabola /parabola (Mersenne Telescope)Document9 pages(A) Parabola /parabola (Mersenne Telescope)Laura PerozaNo ratings yet
- Overdosing PDFDocument13 pagesOverdosing PDFAlida CuevasNo ratings yet
- Nuclear Pore ComplexesDocument9 pagesNuclear Pore ComplexesMNo ratings yet
- Ebbesson, Sven O. E. (1980) - Comparative Neurology of The Telencephalon. The Telencephalon of SnakesDocument39 pagesEbbesson, Sven O. E. (1980) - Comparative Neurology of The Telencephalon. The Telencephalon of SnakesLivia PereiraNo ratings yet
- Zx200-5g ManualDocument13 pagesZx200-5g ManualBeny StephenNo ratings yet
- Garcia-Austt & Patetta-Queirolo 1961aDocument11 pagesGarcia-Austt & Patetta-Queirolo 1961achimera01No ratings yet
- OPT_Manual_2024-01-08Document19 pagesOPT_Manual_2024-01-08丁安磊No ratings yet
- Bergevin 2010Document13 pagesBergevin 2010overkindNo ratings yet
- ZYBIODocument2 pagesZYBIOChristine OlivarNo ratings yet
- 1285 سراجهDocument4 pages1285 سراجهm1.nourianNo ratings yet
- Extracellular Matrix Synthesis by Skeletal Muscle in CultureDocument6 pagesExtracellular Matrix Synthesis by Skeletal Muscle in Cultureranbos6815No ratings yet
- Young's double slit experimentDocument10 pagesYoung's double slit experimentvasea_1No ratings yet
- Molecular Interactions and Fluorescence TechniquesDocument49 pagesMolecular Interactions and Fluorescence TechniquesGuillaumeNo ratings yet
- Gariaev, Peter P._ Chudin, Viktor I._ Komissarov, Gennady G._ Be - SPIE Proceedings [SPIE Optical Memory and Neural Networks - Zvenigorod, Russian Federation (Tuesday 2 April 1991)] Optical Memor (1991, SPIE) [10.1117_12.50Document12 pagesGariaev, Peter P._ Chudin, Viktor I._ Komissarov, Gennady G._ Be - SPIE Proceedings [SPIE Optical Memory and Neural Networks - Zvenigorod, Russian Federation (Tuesday 2 April 1991)] Optical Memor (1991, SPIE) [10.1117_12.50NiklausNo ratings yet
- 2020 Ka MaalDocument11 pages2020 Ka MaalMaharshee RanjanNo ratings yet
- Band Structure And Nuclear Dynamics: Proceedings of the International Conference On Band Structure And Nuclear Dynamics Tulane University, New OrleansFrom EverandBand Structure And Nuclear Dynamics: Proceedings of the International Conference On Band Structure And Nuclear Dynamics Tulane University, New OrleansA.L. GoodmanNo ratings yet
- Total Cross-Section Measurements: Progress in Nuclear PhysicsFrom EverandTotal Cross-Section Measurements: Progress in Nuclear PhysicsNo ratings yet
- Mreznica - 80Document1 pageMreznica - 80AkicaNo ratings yet
- R. M. Shapley and C. Enroth-Cugell: Et AlDocument1 pageR. M. Shapley and C. Enroth-Cugell: Et AlAkicaNo ratings yet
- Mreznica - 73Document1 pageMreznica - 73AkicaNo ratings yet
- Mreznica - 79Document1 pageMreznica - 79AkicaNo ratings yet
- Mreznica - 78Document1 pageMreznica - 78AkicaNo ratings yet
- Visual Adaptation and Retinal Gain ControlsDocument1 pageVisual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- Mreznica - 74Document1 pageMreznica - 74AkicaNo ratings yet
- Visual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)Document1 pageVisual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)AkicaNo ratings yet
- Tpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IDocument1 pageTpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IAkicaNo ratings yet
- Et Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30Document1 pageEt Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30AkicaNo ratings yet
- Mreznica - 64Document1 pageMreznica - 64AkicaNo ratings yet
- Mreznica - 66Document1 pageMreznica - 66AkicaNo ratings yet
- Z,, (Co) Y.L (Co) I, (O ) I, (Co) : Visual Adaptation and Retinal Gain ControlsDocument1 pageZ,, (Co) Y.L (Co) I, (O ) I, (Co) : Visual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- Mreznica - 71Document1 pageMreznica - 71AkicaNo ratings yet
- Mreznica - 61Document1 pageMreznica - 61AkicaNo ratings yet
- 6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oDocument1 page6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oAkicaNo ratings yet
- Time Course of Gain Adjustment in Retinal Ganglion CellsDocument1 pageTime Course of Gain Adjustment in Retinal Ganglion CellsAkicaNo ratings yet
- Do Not: 5. Gain Control in PhotoreceptorsDocument1 pageDo Not: 5. Gain Control in PhotoreceptorsAkicaNo ratings yet
- Mreznica - 58Document1 pageMreznica - 58AkicaNo ratings yet
- Mreznica - 55Document1 pageMreznica - 55AkicaNo ratings yet
- Retina - 53 PDFDocument1 pageRetina - 53 PDFAkicaNo ratings yet
- Retina - 52Document1 pageRetina - 52AkicaNo ratings yet
- Retina - 52Document1 pageRetina - 52AkicaNo ratings yet
- Retina - 52Document1 pageRetina - 52AkicaNo ratings yet
- Visual Adaptation and Retinal Gain ControlsDocument1 pageVisual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- Retina - 51Document1 pageRetina - 51AkicaNo ratings yet
- 3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsDocument1 page3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsAkicaNo ratings yet
- Gizzard Proventriculus Health Marker Avinews Int Sept 22Document11 pagesGizzard Proventriculus Health Marker Avinews Int Sept 22mohamed helmyNo ratings yet
- Animal Trypanosomiasis in Somalia (Paper For Discussion)Document10 pagesAnimal Trypanosomiasis in Somalia (Paper For Discussion)Aamir muse osmanNo ratings yet
- Test Bank For Understanding Pathophysiology 6th Edition by HuetherDocument24 pagesTest Bank For Understanding Pathophysiology 6th Edition by HuetherJulieCooperb25om100% (33)
- Chrysothemis PulchellaDocument2 pagesChrysothemis PulchellaTharayilsijNo ratings yet
- About SharksDocument15 pagesAbout Sharkselias1.1dubeiNo ratings yet
- Blood (Notes)Document12 pagesBlood (Notes)Angel Rose BrillanteNo ratings yet
- Novel components of NAD pathways predictedDocument18 pagesNovel components of NAD pathways predictedGuillermo Domínguez HuertaNo ratings yet
- The Antibacterial and Antifungal Activity of Essential Oil Derived From The Flesh of Nutmeg Fruit 6155Document6 pagesThe Antibacterial and Antifungal Activity of Essential Oil Derived From The Flesh of Nutmeg Fruit 6155Zyuha AiniiNo ratings yet
- Allergy Testing PDFDocument25 pagesAllergy Testing PDFRK MalhotraNo ratings yet
- E CA 72-4 en 9Document3 pagesE CA 72-4 en 9Hassan GillNo ratings yet
- Substance Form and Transprort Learning IssuesDocument8 pagesSubstance Form and Transprort Learning IssuesIftitah N. RNo ratings yet
- Steroid AbuseDocument7 pagesSteroid Abuseapi-400575655No ratings yet
- ProteinsDocument1 pageProteinsKhey AgamNo ratings yet
- Application of "Dispersive Liquid-Liquid Micro Extraction Technique" For The Analysis of Piroxicam in Human Urine and Drug FormulationDocument16 pagesApplication of "Dispersive Liquid-Liquid Micro Extraction Technique" For The Analysis of Piroxicam in Human Urine and Drug FormulationInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Madhu VargDocument36 pagesMadhu VargYash GardhariyaNo ratings yet
- 4d2a1 HD Adequacy Self StudyDocument31 pages4d2a1 HD Adequacy Self StudyWulan SuciNo ratings yet
- Bio 10th - Excretion - Extensive TestDocument1 pageBio 10th - Excretion - Extensive TestDharmendra SinghNo ratings yet
- Pattern Matching: Classifying Organic MoleculesDocument8 pagesPattern Matching: Classifying Organic MoleculesBhaavana AnnaNo ratings yet
- Lecture No 1 CRM 223 DactylosDocument25 pagesLecture No 1 CRM 223 Dactylosjanicaritzpaez45No ratings yet
- Pharmacology - Mechanism of Action of All Drugs - +Document4 pagesPharmacology - Mechanism of Action of All Drugs - +Sahal ShaikhNo ratings yet
- Test 3 (3)Document6 pagesTest 3 (3)Blabla HiNo ratings yet
- Buenas Practicas de Microbiologia USPDocument5 pagesBuenas Practicas de Microbiologia USPGloria Daniela ReyesNo ratings yet
- Infowar and Spiritual Apocalypse The Destiny of Mankind: DR Bill Deagle MDDocument37 pagesInfowar and Spiritual Apocalypse The Destiny of Mankind: DR Bill Deagle MDprateekthaparian100% (1)
- Grade Ix Biology Animal Tissues 2021Document55 pagesGrade Ix Biology Animal Tissues 2021Purvi Prakash KNo ratings yet
- Jeopardy Game 5-Topic Template TaigaDocument54 pagesJeopardy Game 5-Topic Template TaigaDylan SleasmanNo ratings yet
- Package Insert - 06267 - LH - en - 30406 PDFDocument6 pagesPackage Insert - 06267 - LH - en - 30406 PDFadybaila4680No ratings yet
- MD - Abdur - Rahim - MS - Thesis - Genetic Diversity Analysis in Rice (Oryza Sativa)Document10 pagesMD - Abdur - Rahim - MS - Thesis - Genetic Diversity Analysis in Rice (Oryza Sativa)rahimgpbNo ratings yet
- Daun Kersen Penurunan Kadar Kolestrol PDFDocument8 pagesDaun Kersen Penurunan Kadar Kolestrol PDFJumriani HNo ratings yet
- The Five Major Brances of Chemistry and Their Sub-BranchesDocument3 pagesThe Five Major Brances of Chemistry and Their Sub-BranchesYenna Rezano LumbaNo ratings yet
- BPA Decreases Testosterone in Male RatsDocument6 pagesBPA Decreases Testosterone in Male RatsNOVITANo ratings yet























































![Gariaev, Peter P._ Chudin, Viktor I._ Komissarov, Gennady G._ Be - SPIE Proceedings [SPIE Optical Memory and Neural Networks - Zvenigorod, Russian Federation (Tuesday 2 April 1991)] Optical Memor (1991, SPIE) [10.1117_12.50](https://imgv2-1-f.scribdassets.com/img/document/723127591/149x198/4e7160e25d/1713164131?v=1)