You might also like
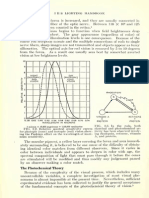
- 0"4 I ' 0"2 4 Log I 2 Photopic TrofandsDocument1 page0"4 I ' 0"2 4 Log I 2 Photopic TrofandsAkicaNo ratings yet
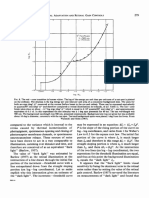
- 2.1.1.1. Aguilar and Stiles' Experiment.: Visual Adaptation and Retinal G A I N Controls 2 7 7Document1 page2.1.1.1. Aguilar and Stiles' Experiment.: Visual Adaptation and Retinal G A I N Controls 2 7 7AkicaNo ratings yet
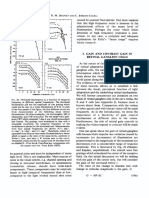
- Visual adaptation and retinal gain controlsDocument1 pageVisual adaptation and retinal gain controlsAkicaNo ratings yet
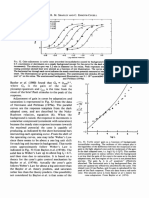
- Retina - 43Document1 pageRetina - 43AkicaNo ratings yet
- Mreznica - 59Document1 pageMreznica - 59AkicaNo ratings yet
- Retina - 24Document1 pageRetina - 24AkicaNo ratings yet
- I Ooo" ': IC) A ", I (D)Document1 pageI Ooo" ': IC) A ", I (D)AkicaNo ratings yet
- The Vibrational Spectra of Phenol and phenol-ODDocument11 pagesThe Vibrational Spectra of Phenol and phenol-ODMaulida Azizza ShizenNo ratings yet
- University of California Los Angeles .. - ,,-. - .: Approved For Release Cia-Rdp96-007B7R0004000Booic RDDocument6 pagesUniversity of California Los Angeles .. - ,,-. - .: Approved For Release Cia-Rdp96-007B7R0004000Booic RDChadNo ratings yet
- R - M - Shapley and C - Enroth-CugellDocument1 pageR - M - Shapley and C - Enroth-CugellAkicaNo ratings yet
- Tpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IDocument1 pageTpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IAkicaNo ratings yet
- Pbse BandstrucureDocument12 pagesPbse BandstrucurevinaychimalgiNo ratings yet
- Retina - 40Document1 pageRetina - 40AkicaNo ratings yet
- Abraham1988 PDFDocument4 pagesAbraham1988 PDFSRS 2016No ratings yet
- ApplOpt 38 1657 1999 MatrixInversionRetrievalDocument6 pagesApplOpt 38 1657 1999 MatrixInversionRetrievalZhao WenwenNo ratings yet
- Diffuse Reflectance Spectra and Optical Properties of Some Iron and Titanium Oxides and OxyhydroxidesDocument8 pagesDiffuse Reflectance Spectra and Optical Properties of Some Iron and Titanium Oxides and Oxyhydroxidesicpsdt.cuetNo ratings yet
- Ch17 Ch18 Review QuestionsDocument38 pagesCh17 Ch18 Review QuestionsLuis Rodriguez0% (1)
- Breaking Abbe's Diffraction Resolution Limit in Fluorescence MicrosDocument9 pagesBreaking Abbe's Diffraction Resolution Limit in Fluorescence Microspál locNo ratings yet
- OF of Stellar Spectra: Application DicyaninDocument25 pagesOF of Stellar Spectra: Application DicyaninLevicano100% (1)
- Rioja 1977Document5 pagesRioja 1977hcmadhuNo ratings yet
- Mreznica - 61Document1 pageMreznica - 61AkicaNo ratings yet
- The Human Visual System and Perception: / (A,) Pea,) L (A,)Document24 pagesThe Human Visual System and Perception: / (A,) Pea,) L (A,)kaiser yasserNo ratings yet
- Longtransient Effects in Lasers With Inserted Liquid SamplesDocument7 pagesLongtransient Effects in Lasers With Inserted Liquid SamplesDarwin GuangaNo ratings yet
- Research Department Tannoy Velocity Microphone: Report No. M.007 Serial NoDocument5 pagesResearch Department Tannoy Velocity Microphone: Report No. M.007 Serial NoalpNo ratings yet
- Retina 11Document1 pageRetina 11AkicaNo ratings yet
- Retina - 35Document1 pageRetina - 35AkicaNo ratings yet
- 1.reristivity Logging ToolDocument10 pages1.reristivity Logging ToolFatimah AzzahraNo ratings yet
- Fluctuations of Starlight and SkylightDocument26 pagesFluctuations of Starlight and Skylightskr2010No ratings yet
- Telecont: Single-Photon Counter ForDocument7 pagesTelecont: Single-Photon Counter ForDaliaBarrancoNo ratings yet
- Retina - 18Document1 pageRetina - 18AkicaNo ratings yet
- Maekawa1968 PDFDocument17 pagesMaekawa1968 PDFMarely Teresa Ruiz AntolinezNo ratings yet
- BF00576726Document2 pagesBF00576726AZIL KenzaNo ratings yet
- Confort Ingles 23Document1 pageConfort Ingles 23jade485No ratings yet
- 03 14 PerronDocument12 pages03 14 Perronlucille.bonnierNo ratings yet
- Base SchiffDocument10 pagesBase SchiffDánica Nicoll Rojas MolinaNo ratings yet
- Near-IR Spectroscopy Determines Epoxy Resin CureDocument11 pagesNear-IR Spectroscopy Determines Epoxy Resin CureRick MortyNo ratings yet
- Visual Adaptation and Retinal Gain ControlsDocument1 pageVisual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- Facts and Artifacts in Near-Field Optical Microscopy: Related ArticlesDocument8 pagesFacts and Artifacts in Near-Field Optical Microscopy: Related ArticlesClaudio BiaginiNo ratings yet
- PhysRevB.66.195415Document9 pagesPhysRevB.66.195415CH YNo ratings yet
- Prinsip HuygensDocument36 pagesPrinsip Huygensqc12345No ratings yet
- Colored InversionDocument5 pagesColored Inversiondanial bramantaNo ratings yet
- O / e (AT o'A'T) : Log SDocument1 pageO / e (AT o'A'T) : Log SAkicaNo ratings yet
- C. Enroth-Cugell: Et Al.Document1 pageC. Enroth-Cugell: Et Al.AkicaNo ratings yet
- Types of Fluorescence Transitions in Atomic Fluorescence SpectrometryDocument3 pagesTypes of Fluorescence Transitions in Atomic Fluorescence SpectrometryAimen KhalidNo ratings yet
- B.Eng - Sc. (Mechanical Engineering), Universityof Alexandria, 1970 M.Eng - Sc. (Mechanical Engineering), Universityof Alexandria, 1974Document143 pagesB.Eng - Sc. (Mechanical Engineering), Universityof Alexandria, 1970 M.Eng - Sc. (Mechanical Engineering), Universityof Alexandria, 1974Samik MaitiNo ratings yet
- Blackshear 1956 NACA The Growth of Disturbances in A Flame Generated Shear RegionDocument11 pagesBlackshear 1956 NACA The Growth of Disturbances in A Flame Generated Shear RegionPrem ChandNo ratings yet
- Serial 1949/29:, Inv) ! Stigatlon of Belmhoitz Resouators Report No. B.041Document26 pagesSerial 1949/29:, Inv) ! Stigatlon of Belmhoitz Resouators Report No. B.041AntonioPalloneNo ratings yet
- Mouth State Detection Using Log-Polar SignaturesDocument11 pagesMouth State Detection Using Log-Polar SignaturesMohammad AkifNo ratings yet
- Am. J - Med. and Hyg.,: Communications Enr VolDocument3 pagesAm. J - Med. and Hyg.,: Communications Enr VolFELIPE DANIEL MONTERO BRUNINo ratings yet
- Jresv70cn2p75 A1bDocument14 pagesJresv70cn2p75 A1bZé Pontes DetCordNo ratings yet
- The Effect of Systox On Ionic Fluxes in Excised Barley RootsDocument10 pagesThe Effect of Systox On Ionic Fluxes in Excised Barley RootsSh1vaNo ratings yet
- Et Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30Document1 pageEt Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30AkicaNo ratings yet
- R. M. Shapley and C. Enroth-Cugell: Range AnDocument1 pageR. M. Shapley and C. Enroth-Cugell: Range AnAkicaNo ratings yet
- E Lighting: BetweenDocument1 pageE Lighting: BetweenreacharunkNo ratings yet
- bose1997_Preparation_of_nonclassical_states_in_cavities_with_a_movingDocument12 pagesbose1997_Preparation_of_nonclassical_states_in_cavities_with_a_movingVeronika TreumovaNo ratings yet
- Metformin and Sitagliptin Prolonged-Release TabletsDocument7 pagesMetformin and Sitagliptin Prolonged-Release Tabletsmykiend2002No ratings yet
- Night MiopiaDocument8 pagesNight MiopiamurilobsouzaNo ratings yet
- Oth R MetalDocument10 pagesOth R MetalBhavesh ShimpiNo ratings yet
- Medical Division, Air Force Systems Commnand, e 62F Wright-Patterson Air Force Base, Ohio 45433i210-7 78-45 Feb 8Document15 pagesMedical Division, Air Force Systems Commnand, e 62F Wright-Patterson Air Force Base, Ohio 45433i210-7 78-45 Feb 8Bryan396No ratings yet
- Mreznica - 78Document1 pageMreznica - 78AkicaNo ratings yet
- Mreznica - 80Document1 pageMreznica - 80AkicaNo ratings yet
- Mreznica - 79Document1 pageMreznica - 79AkicaNo ratings yet
- Mreznica - 71Document1 pageMreznica - 71AkicaNo ratings yet
- Visual Adaptation and Retinal Gain ControlsDocument1 pageVisual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- 6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oDocument1 page6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oAkicaNo ratings yet
- Z,, (Co) Y.L (Co) I, (O ) I, (Co) : Visual Adaptation and Retinal Gain ControlsDocument1 pageZ,, (Co) Y.L (Co) I, (O ) I, (Co) : Visual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- R. M. Shapley and C. Enroth-Cugell: Et AlDocument1 pageR. M. Shapley and C. Enroth-Cugell: Et AlAkicaNo ratings yet
- Et Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30Document1 pageEt Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30AkicaNo ratings yet
- Mreznica - 74Document1 pageMreznica - 74AkicaNo ratings yet
- Visual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)Document1 pageVisual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)AkicaNo ratings yet
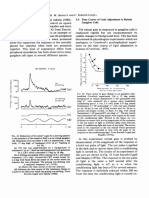
- Time Course of Gain Adjustment in Retinal Ganglion CellsDocument1 pageTime Course of Gain Adjustment in Retinal Ganglion CellsAkicaNo ratings yet
- Mreznica - 66Document1 pageMreznica - 66AkicaNo ratings yet
- Mreznica - 73Document1 pageMreznica - 73AkicaNo ratings yet
- Mreznica - 64Document1 pageMreznica - 64AkicaNo ratings yet
- Do Not: 5. Gain Control in PhotoreceptorsDocument1 pageDo Not: 5. Gain Control in PhotoreceptorsAkicaNo ratings yet
- Tpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IDocument1 pageTpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IAkicaNo ratings yet
- Mreznica - 58Document1 pageMreznica - 58AkicaNo ratings yet
- Mreznica - 60Document1 pageMreznica - 60AkicaNo ratings yet
- Mreznica - 61Document1 pageMreznica - 61AkicaNo ratings yet
- Retina - 52Document1 pageRetina - 52AkicaNo ratings yet
- Mreznica - 55Document1 pageMreznica - 55AkicaNo ratings yet
- 4.3. Horizontal Cells: F I I FDocument1 page4.3. Horizontal Cells: F I I FAkicaNo ratings yet
- Visual Adaptation and Retinal Gain ControlsDocument1 pageVisual Adaptation and Retinal Gain ControlsAkicaNo ratings yet
- Retina - 52Document1 pageRetina - 52AkicaNo ratings yet
- Retina - 51Document1 pageRetina - 51AkicaNo ratings yet
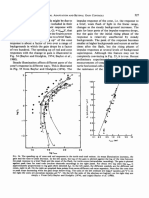
- 3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsDocument1 page3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsAkicaNo ratings yet
- Retina - 53 PDFDocument1 pageRetina - 53 PDFAkicaNo ratings yet
- Retina - 52Document1 pageRetina - 52AkicaNo ratings yet
- Chicken Foot DissectionDocument2 pagesChicken Foot DissectionGacostaza09No ratings yet
- KS4 Basic Life Support PresentationDocument29 pagesKS4 Basic Life Support PresentationDixiline MarzoNo ratings yet
- Rhodophyta Fact SheetDocument3 pagesRhodophyta Fact Sheetapi-299846771No ratings yet
- Pex 09 03Document4 pagesPex 09 03Marcela Anco Sotomayor50% (4)
- Answer Questions5 1Document3 pagesAnswer Questions5 1yo-chengNo ratings yet
- Khalid Complete Anesthesia Notes PDFDocument361 pagesKhalid Complete Anesthesia Notes PDFDrMuhammad Ishfaq Habib100% (3)
- 2 Quarter Examination S.Y. 2019-2020: Earth Life and Science - Grade 11Document6 pages2 Quarter Examination S.Y. 2019-2020: Earth Life and Science - Grade 11Mihatsu TakiNo ratings yet
- What Is NeurobicsDocument6 pagesWhat Is Neurobicsvarun_009No ratings yet
- Pulsus Paradoxus - Wikip PDFDocument4 pagesPulsus Paradoxus - Wikip PDFAniket MittalNo ratings yet
- Medical Surgical Nursing Exam 19 NLE Style 100 Items Nurseslabs December 2012 NLE ResultsDocument21 pagesMedical Surgical Nursing Exam 19 NLE Style 100 Items Nurseslabs December 2012 NLE ResultsKaren Ann M. Estrada100% (1)
- Fundamentals of Anatomy and Physiology 8Th Edition Martini Test Bank Full Chapter PDFDocument50 pagesFundamentals of Anatomy and Physiology 8Th Edition Martini Test Bank Full Chapter PDFgillanolympia4091253100% (9)
- New Estrogen and ProgesteroneDocument39 pagesNew Estrogen and ProgesteroneWegrimel AriegaraNo ratings yet
- Neonatal Jaundice: Prevention, Assessment and Management 2Document20 pagesNeonatal Jaundice: Prevention, Assessment and Management 2kyawswakyawswaNo ratings yet
- Tratamiento Quirúrgico Del Mal Perforante Plantar: C.M. Hernández-Cañete, J. Borroto-PachecoDocument7 pagesTratamiento Quirúrgico Del Mal Perforante Plantar: C.M. Hernández-Cañete, J. Borroto-PachecoDiegoNo ratings yet
- Physiology Course: Dr. Velu M. RachelDocument40 pagesPhysiology Course: Dr. Velu M. RachelKunda JosephNo ratings yet
- Nursing Care PlanDocument7 pagesNursing Care Planmcd7r883% (6)
- Screenshot 2021-06-23 at 11.36.48 AMDocument116 pagesScreenshot 2021-06-23 at 11.36.48 AMSmitha ShekarNo ratings yet
- Glycine MetabolismDocument28 pagesGlycine MetabolismSuryansh GoyalNo ratings yet
- TRIO For Flute, Guitar, and CelloDocument44 pagesTRIO For Flute, Guitar, and CelloNick Schuller100% (1)
- Unstable Angina With Underlying DyslipidaemiaDocument9 pagesUnstable Angina With Underlying DyslipidaemiaAiman ArifinNo ratings yet
- Plant Body Plant Body: Why Is Histology Important?Document6 pagesPlant Body Plant Body: Why Is Histology Important?ZyreeneNicoleNo ratings yet
- Physical Education Nine Ten English VersionDocument149 pagesPhysical Education Nine Ten English Versiondsafg87g87gawdwNo ratings yet
- Callus CultureDocument24 pagesCallus CultureHabiba Majeed MalikNo ratings yet
- Nursing Process in The Care of The Preterm InfantDocument5 pagesNursing Process in The Care of The Preterm InfantizwanNo ratings yet
- Manage Upper GI Bleed in 33yo Hotel ReceptionistDocument42 pagesManage Upper GI Bleed in 33yo Hotel ReceptionistMuhammad Naveed AslamNo ratings yet
- Psychology Canadian 2nd Edition Ciccarelli Test BankDocument42 pagesPsychology Canadian 2nd Edition Ciccarelli Test BankMichaelAllenrazne100% (10)
- 41 Emotions As Expressed Through Body LanguageDocument9 pages41 Emotions As Expressed Through Body LanguageSocaciu Gabriel100% (1)
- Nursing Care Plan For Postpartum HemorrhageDocument2 pagesNursing Care Plan For Postpartum HemorrhageDianne Mae100% (1)
- Thermoregulation in PiscesDocument4 pagesThermoregulation in PiscesAbellya Nella Morte100% (3)
- All About SharksDocument3 pagesAll About SharksGutmonarchNo ratings yet
- Quantum Spirituality: Science, Gnostic Mysticism, and Connecting with Source ConsciousnessFrom EverandQuantum Spirituality: Science, Gnostic Mysticism, and Connecting with Source ConsciousnessRating: 4 out of 5 stars4/5 (6)
- A Brief History of Time: From the Big Bang to Black HolesFrom EverandA Brief History of Time: From the Big Bang to Black HolesRating: 4 out of 5 stars4/5 (2193)
- Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseFrom EverandDark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseRating: 3.5 out of 5 stars3.5/5 (69)
- Summary and Interpretation of Reality TransurfingFrom EverandSummary and Interpretation of Reality TransurfingRating: 5 out of 5 stars5/5 (5)
- Strange Angel: The Otherworldly Life of Rocket Scientist John Whiteside ParsonsFrom EverandStrange Angel: The Otherworldly Life of Rocket Scientist John Whiteside ParsonsRating: 4 out of 5 stars4/5 (94)
- A Beginner's Guide to Constructing the Universe: The Mathematical Archetypes of Nature, Art, and ScienceFrom EverandA Beginner's Guide to Constructing the Universe: The Mathematical Archetypes of Nature, Art, and ScienceRating: 4 out of 5 stars4/5 (51)
- The Beginning of Infinity: Explanations That Transform the WorldFrom EverandThe Beginning of Infinity: Explanations That Transform the WorldRating: 5 out of 5 stars5/5 (60)
- Packing for Mars: The Curious Science of Life in the VoidFrom EverandPacking for Mars: The Curious Science of Life in the VoidRating: 4 out of 5 stars4/5 (1395)
- Knocking on Heaven's Door: How Physics and Scientific Thinking Illuminate the Universe and the Modern WorldFrom EverandKnocking on Heaven's Door: How Physics and Scientific Thinking Illuminate the Universe and the Modern WorldRating: 3.5 out of 5 stars3.5/5 (64)
- Lost in Math: How Beauty Leads Physics AstrayFrom EverandLost in Math: How Beauty Leads Physics AstrayRating: 4.5 out of 5 stars4.5/5 (125)
- The Tao of Physics: An Exploration of the Parallels between Modern Physics and Eastern MysticismFrom EverandThe Tao of Physics: An Exploration of the Parallels between Modern Physics and Eastern MysticismRating: 4 out of 5 stars4/5 (500)
- The Physics of God: How the Deepest Theories of Science Explain Religion and How the Deepest Truths of Religion Explain ScienceFrom EverandThe Physics of God: How the Deepest Theories of Science Explain Religion and How the Deepest Truths of Religion Explain ScienceRating: 4.5 out of 5 stars4.5/5 (23)
- The End of Everything: (Astrophysically Speaking)From EverandThe End of Everything: (Astrophysically Speaking)Rating: 4.5 out of 5 stars4.5/5 (157)
- Quantum Physics: What Everyone Needs to KnowFrom EverandQuantum Physics: What Everyone Needs to KnowRating: 4.5 out of 5 stars4.5/5 (48)
- Quantum Physics for Beginners Who Flunked Math And Science: Quantum Mechanics And Physics Made Easy Guide In Plain Simple EnglishFrom EverandQuantum Physics for Beginners Who Flunked Math And Science: Quantum Mechanics And Physics Made Easy Guide In Plain Simple EnglishRating: 4.5 out of 5 stars4.5/5 (18)
- Bedeviled: A Shadow History of Demons in ScienceFrom EverandBedeviled: A Shadow History of Demons in ScienceRating: 5 out of 5 stars5/5 (5)
- Midnight in Chernobyl: The Story of the World's Greatest Nuclear DisasterFrom EverandMidnight in Chernobyl: The Story of the World's Greatest Nuclear DisasterRating: 4.5 out of 5 stars4.5/5 (409)
- Black Holes: The Key to Understanding the UniverseFrom EverandBlack Holes: The Key to Understanding the UniverseRating: 4.5 out of 5 stars4.5/5 (13)
- In Search of Schrödinger’s Cat: Quantum Physics and RealityFrom EverandIn Search of Schrödinger’s Cat: Quantum Physics and RealityRating: 4 out of 5 stars4/5 (380)
- The Sounds of Life: How Digital Technology Is Bringing Us Closer to the Worlds of Animals and PlantsFrom EverandThe Sounds of Life: How Digital Technology Is Bringing Us Closer to the Worlds of Animals and PlantsRating: 5 out of 5 stars5/5 (5)
- The Simulated Multiverse: An MIT Computer Scientist Explores Parallel Universes, The Simulation Hypothesis, Quantum Computing and the Mandela EffectFrom EverandThe Simulated Multiverse: An MIT Computer Scientist Explores Parallel Universes, The Simulation Hypothesis, Quantum Computing and the Mandela EffectRating: 4.5 out of 5 stars4.5/5 (20)