You might also like
- Media 823904 SMXXDocument26 pagesMedia 823904 SMXXДемьян МатченкоNo ratings yet
- 10 1016@j BJ 2019 10 005Document10 pages10 1016@j BJ 2019 10 005hiwmacrigeeeNo ratings yet
- SARS-CoV-2 Variants - UK + South African + Brazil Variants - Key AtfDocument1 pageSARS-CoV-2 Variants - UK + South African + Brazil Variants - Key AtfJuan Manuel Tapia AlzateNo ratings yet
- Overview - Regulation of ApoptosisDocument1 pageOverview - Regulation of ApoptosisAym mantezNo ratings yet
- A CRISPR Path To Cutting-Edge MaterialsDocument4 pagesA CRISPR Path To Cutting-Edge MaterialsGabriela ArvelaezNo ratings yet
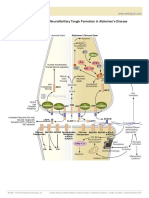
- Amyloid Plaque and Neurofibrillary Tangle Formation in Alzheimer's DiseaseDocument2 pagesAmyloid Plaque and Neurofibrillary Tangle Formation in Alzheimer's DiseaseMelanieNo ratings yet
- CRISPRDocument17 pagesCRISPRKiran MishraNo ratings yet
- Crispr-Cas12B Enables Efficient Plant Genome Engineering: Brief CommunicationDocument9 pagesCrispr-Cas12B Enables Efficient Plant Genome Engineering: Brief CommunicationIram ExpósitoNo ratings yet
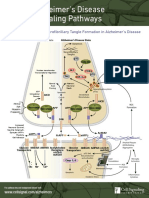
- Https - Learn - Cellsignal.com - Hubfs - Pdfs - 15 Fly Alzh Ref m096 Eng 00 Alzheimers PW Handout DigitalDocument2 pagesHttps - Learn - Cellsignal.com - Hubfs - Pdfs - 15 Fly Alzh Ref m096 Eng 00 Alzheimers PW Handout DigitaltamaraNo ratings yet
- Worksheet16 Transcription To TranslationDocument3 pagesWorksheet16 Transcription To Translationliterally deadNo ratings yet
- The Next Generation of CRISPR-Cas Technologies and ApplicationsDocument18 pagesThe Next Generation of CRISPR-Cas Technologies and ApplicationsCursos HMMNo ratings yet
- CRISPR Knockouts GuideDocument20 pagesCRISPR Knockouts GuideVương HoàngNo ratings yet
- CRISPR in NeuroscienceDocument9 pagesCRISPR in NeuroscienceVlad TomaNo ratings yet
- CRISPR Guide: Compare Types II, V, VI SystemsDocument1 pageCRISPR Guide: Compare Types II, V, VI SystemsLaila VazNo ratings yet
- ViruzhinkiDocument20 pagesViruzhinkiЕгор ЧудовскийNo ratings yet
- 10.1038@s41586 019 0908 XDocument23 pages10.1038@s41586 019 0908 XJ ENo ratings yet
- METTL16 Antagonizes MRE11-mediated DNA End Resection and Confers Synthetic Lethality To PARP Inhibition in Pancreatic Ductal AdenocarcinomaDocument31 pagesMETTL16 Antagonizes MRE11-mediated DNA End Resection and Confers Synthetic Lethality To PARP Inhibition in Pancreatic Ductal AdenocarcinomaJackson AlbertNo ratings yet
- Crispr Cas-9 Project SummaryDocument3 pagesCrispr Cas-9 Project SummaryKavya DholariyaNo ratings yet
- Lab Report DNA PDFDocument8 pagesLab Report DNA PDFpadhiyararpitaNo ratings yet
- 3 ViraDocument9 pages3 ViraVira AlindaNo ratings yet
- 调控 CRISPR-Cas9 系统用于基因编辑的研究进展: Acta Chimica SinicaDocument8 pages调控 CRISPR-Cas9 系统用于基因编辑的研究进展: Acta Chimica SinicaChou CharlesNo ratings yet
- 调控 CRISPR-Cas9 系统用于基因编辑的研究进展: Acta Chimica SinicaDocument8 pages调控 CRISPR-Cas9 系统用于基因编辑的研究进展: Acta Chimica SinicaChou CharlesNo ratings yet
- ACFrOgDWGR7iXyIO5GsdkRtH3Exx5VlMTm1ltALd8ms37CeVjv9YaiNPOHzcK0nUC FX8qH3sSuYeYEltAe KuPE1xyq6MOkpGx9ZKKbFui3xpyDepIk93 KGP VRsZIHpQNt8p4vnTrgvUzujsTDocument9 pagesACFrOgDWGR7iXyIO5GsdkRtH3Exx5VlMTm1ltALd8ms37CeVjv9YaiNPOHzcK0nUC FX8qH3sSuYeYEltAe KuPE1xyq6MOkpGx9ZKKbFui3xpyDepIk93 KGP VRsZIHpQNt8p4vnTrgvUzujsTAniket AmanNo ratings yet
- Cas 3Document1 pageCas 3api-559521963No ratings yet
- English Grammar PDF Study Material by Angel AcademyDocument116 pagesEnglish Grammar PDF Study Material by Angel AcademyVijay VarsaniNo ratings yet
- Foreign DNA Capture During CRISPR-Cas Immunity Doudna 2015Document13 pagesForeign DNA Capture During CRISPR-Cas Immunity Doudna 2015Marina Célia Nunes Ferreira Da C SilvaNo ratings yet
- fpls-13-839001Document17 pagesfpls-13-839001xixew84371No ratings yet
- Global Profiling of RNA-binding Protein TargetDocument28 pagesGlobal Profiling of RNA-binding Protein TargetYanjiaoNo ratings yet
- ใบงานสัปดาห์ที่ 3-4Document2 pagesใบงานสัปดาห์ที่ 3-4WEERAWAT SANGPECHNo ratings yet
- ScienceBuddies CRISPR BrainstormingWorksheet AnswerKeyDocument4 pagesScienceBuddies CRISPR BrainstormingWorksheet AnswerKeyJuan Sebastian CeleitaNo ratings yet
- KEY Resources TableDocument6 pagesKEY Resources TableDNo ratings yet
- Molecular Biology Workflow Solutions BrochureDocument62 pagesMolecular Biology Workflow Solutions BrochureJoséMaríaMoralesMuñozNo ratings yet
- 12 Laboratory Diagnosis of HIV 2Document11 pages12 Laboratory Diagnosis of HIV 2Melody Jane PardilloNo ratings yet
- Anticuerpos PDFDocument1 pageAnticuerpos PDFOmar Enrique Ocaña GarciaNo ratings yet
- HDI Design Part 1Document103 pagesHDI Design Part 1the_citizen89No ratings yet
- Biomolecules: Recent Advances in CRISPRDocument16 pagesBiomolecules: Recent Advances in CRISPRDouglas SouzaNo ratings yet
- Build A Cell Card Game WorksheetDocument2 pagesBuild A Cell Card Game WorksheetSu AlghNo ratings yet
- Build A Cell Card Game WorksheetDocument2 pagesBuild A Cell Card Game WorksheetlibrahimliNo ratings yet
- Engineering Complex Synthetic Transcriptional Programs with CRISPR RNA ScaffoldsDocument12 pagesEngineering Complex Synthetic Transcriptional Programs with CRISPR RNA ScaffoldsLetícia AlibertiNo ratings yet
- Repair of Double-Strand Breaks Induced by Crispr-Cas9 Leads To Large Deletions and Complex RearrangementsDocument10 pagesRepair of Double-Strand Breaks Induced by Crispr-Cas9 Leads To Large Deletions and Complex RearrangementsMarlon Bundalian CantalNo ratings yet
- Analyzing Snork DNA and TraitsDocument3 pagesAnalyzing Snork DNA and TraitsBrian NguyenNo ratings yet
- CHAPTER 5 - EukaryoticDocument2 pagesCHAPTER 5 - EukaryoticYuume YuuNo ratings yet
- Serotonin and Its Role in PsychiatryDocument46 pagesSerotonin and Its Role in PsychiatryAditya AgrawalNo ratings yet
- Pro To ZoologyDocument9 pagesPro To Zoologytxhj82xcnmNo ratings yet
- English Grammar PDF DownloadDocument198 pagesEnglish Grammar PDF DownloadAshish ShahNo ratings yet
- Transcription )Document5 pagesTranscription )Kotchaphan ChansawangNo ratings yet
- _Genome editing and crispr cas 9Document12 pages_Genome editing and crispr cas 9HemsaiYadavNo ratings yet
- Chenqi Liu - 11 - DNA Vs RNA and Protein Synthesis - Amoeba Sisters - SHODocument2 pagesChenqi Liu - 11 - DNA Vs RNA and Protein Synthesis - Amoeba Sisters - SHOthomasmarteauhNo ratings yet
- Molecular Biology Workflow Solutions BrochureDocument56 pagesMolecular Biology Workflow Solutions BrochureBishoy F. YoussefNo ratings yet
- Increasing the Specificity of CRISPR Systems With Engineered RNA Secondary StructuresDocument14 pagesIncreasing the Specificity of CRISPR Systems With Engineered RNA Secondary StructuresrmbichiNo ratings yet
- Virus RNA BicatenarioDocument4 pagesVirus RNA BicatenarioXavier Munayco OrtizNo ratings yet
- Crispr Poster WebDocument1 pageCrispr Poster Webzero431No ratings yet
- Gabr Mahmoud ThesisDocument35 pagesGabr Mahmoud Thesismahmoud gabrNo ratings yet
- Cell Division, Cell Cycle and Cell TransportDocument3 pagesCell Division, Cell Cycle and Cell Transportandrea.baceovaNo ratings yet
- CRISPR InfographicDocument1 pageCRISPR InfographicSantino Dev Joseph Xavier (Sji)No ratings yet
- Quiz DnaDocument8 pagesQuiz DnaFriska Aprianti100% (3)
- Apoptosis Poster March 20106547Document1 pageApoptosis Poster March 20106547Achmad JunaidiNo ratings yet
- Cas12a ChRDNA - A Novel Approach To Gene-Editing TherapyDocument5 pagesCas12a ChRDNA - A Novel Approach To Gene-Editing TherapyBîkes NâçarNo ratings yet
- The Gut Microbiota-Brain AxisDocument15 pagesThe Gut Microbiota-Brain AxisMateo MendozaNo ratings yet
- 2020-Review-CRISPR Screens in MicrobiomesDocument8 pages2020-Review-CRISPR Screens in MicrobiomesCristian Felipe Sandoval QuiñonezNo ratings yet
- 2020-Review-Analysis of Microbial CompositionsDocument13 pages2020-Review-Analysis of Microbial CompositionsCristian Felipe Sandoval QuiñonezNo ratings yet
- 2020-Review-A Systems Approach To Infectious DiseaseDocument16 pages2020-Review-A Systems Approach To Infectious DiseaseCristian Felipe Sandoval QuiñonezNo ratings yet
- Actividad 4 Artículo 2Document11 pagesActividad 4 Artículo 2byronNo ratings yet
- 2019-Review-The Microbiome and AgingDocument23 pages2019-Review-The Microbiome and AgingCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Integrating Systems and Synthetic Biology To Understand and Engineer MicrobiomesDocument35 pages2021-Integrating Systems and Synthetic Biology To Understand and Engineer MicrobiomesCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Gut Microbiota in Cancer Immune Response and ImmunotherapyDocument14 pages2021-Review-Gut Microbiota in Cancer Immune Response and ImmunotherapyCristian Felipe Sandoval QuiñonezNo ratings yet
- The Microbiome, Cancer, and Cancer Therapy PDFDocument12 pagesThe Microbiome, Cancer, and Cancer Therapy PDFTRAECY LORENA BROCKWAY CLAVERÍANo ratings yet
- 2021 Review Microbiome and CancerDocument25 pages2021 Review Microbiome and CancerCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Engineering The MicrobiomeDocument21 pages2021-Review-Engineering The MicrobiomeCristian Felipe Sandoval QuiñonezNo ratings yet
- 2020-Review-Population Genetics in The Human MicrobiomeDocument15 pages2020-Review-Population Genetics in The Human MicrobiomeCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Mobilome and MicrobiomeDocument13 pages2021-Review-Mobilome and MicrobiomeCristian Felipe Sandoval QuiñonezNo ratings yet
- 2020-Review-Zebrafish Microbiome Studies Make WavesDocument7 pages2020-Review-Zebrafish Microbiome Studies Make WavesCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Phages To Modulate The Microbiome and ImmunityDocument16 pages2021-Review-Phages To Modulate The Microbiome and ImmunityCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Microbiota Effects On CarcinogenesisDocument19 pages2021-Review-Microbiota Effects On CarcinogenesisCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-CRISPR Technologies and PAM-free NucleaseDocument12 pages2021-Review-CRISPR Technologies and PAM-free NucleaseCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Organoids and Organs-On-Chips Gut-Microbe InteractionsDocument12 pages2021-Review-Organoids and Organs-On-Chips Gut-Microbe InteractionsCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-The Microbiome and Human CancerDocument15 pages2021-Review-The Microbiome and Human CancerCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Tweak To Treat Reprograming Bacteria For Cancer TreatmentDocument18 pages2021-Review-Tweak To Treat Reprograming Bacteria For Cancer TreatmentCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-New Technologies For Developing Phage-Based Tools To Manipulate The Human MicrobiomeDocument12 pages2021-Review-New Technologies For Developing Phage-Based Tools To Manipulate The Human MicrobiomeCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Anti-CRISPRs Go Viral The Infection Biology of CRISPR-Cas InhibitorsDocument11 pages2021-Review-Anti-CRISPRs Go Viral The Infection Biology of CRISPR-Cas InhibitorsCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Tumour Neoantigen Mimicry by Microbial Species in Cancer ImmunotherapyDocument11 pages2021-Review-Tumour Neoantigen Mimicry by Microbial Species in Cancer ImmunotherapyCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Controlling and Enhancing CRISPRDocument10 pages2021-Review-Controlling and Enhancing CRISPRCristian Felipe Sandoval QuiñonezNo ratings yet
- 2020-Structures and Strategies of Anti-CRISPR-Mediated Immune SuppressionDocument17 pages2020-Structures and Strategies of Anti-CRISPR-Mediated Immune SuppressionCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Targeting The Gut Microbiome To Mitigate Immunotherapy-Induced Colitis in CancerDocument11 pages2021-Review-Targeting The Gut Microbiome To Mitigate Immunotherapy-Induced Colitis in CancerCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-The Human Virome Assembly, Composition and Host InteractionsDocument14 pages2021-Review-The Human Virome Assembly, Composition and Host InteractionsCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Chemical and Optical Control of CRISPRDocument9 pages2021-Review-Chemical and Optical Control of CRISPRCristian Felipe Sandoval QuiñonezNo ratings yet
- 2020-Review-Technologies and Computational Analysis Strategies For CRISPR ApplicationsDocument19 pages2020-Review-Technologies and Computational Analysis Strategies For CRISPR ApplicationsCristian Felipe Sandoval QuiñonezNo ratings yet
- 2021-Review-Transcriptional Silencers Driving Gene Expression With The Brakes OnDocument14 pages2021-Review-Transcriptional Silencers Driving Gene Expression With The Brakes OnCristian Felipe Sandoval QuiñonezNo ratings yet
- LAS 4 - Recognize The Major Categories of Biomolecules Such As Carbohydrates, Lipids, Proteins, and Nucleic AcidsDocument3 pagesLAS 4 - Recognize The Major Categories of Biomolecules Such As Carbohydrates, Lipids, Proteins, and Nucleic AcidsSalve SerranoNo ratings yet
- Ganong's Physiology Chapter 1-2 ReviewDocument2 pagesGanong's Physiology Chapter 1-2 ReviewMaine AsuncionNo ratings yet
- 2011 고분자 - 분야 - 심사실무가이드Document126 pages2011 고분자 - 분야 - 심사실무가이드ChinsuW.LeeNo ratings yet
- Cytology CytoskeletonDocument48 pagesCytology CytoskeletonMitzel Sapalo0% (1)
- Amoeba Sisters Video Recap:BiomoleculesDocument2 pagesAmoeba Sisters Video Recap:BiomoleculesKimo GamerYTNo ratings yet
- Genetics From Genes To Genomes 5th Edition by Hartwell Goldberg Fischer ISBN Solution ManualDocument6 pagesGenetics From Genes To Genomes 5th Edition by Hartwell Goldberg Fischer ISBN Solution Manualrose100% (27)
- Biological Molecules: Mrs. Mary Vienne M. PascualDocument73 pagesBiological Molecules: Mrs. Mary Vienne M. PascualVienne Monroid100% (1)
- 5 HemostasisDocument35 pages5 Hemostasissamar yousif mohamedNo ratings yet
- P GEXmapDocument1 pageP GEXmapAnonymous byoeVONo ratings yet
- Mapas de relaciones competencialesDocument48 pagesMapas de relaciones competencialesPilar MajanoNo ratings yet
- PCR Lab ReportDocument16 pagesPCR Lab ReportAdibah Ahmad100% (1)
- Recombinant DNA TechnologyDocument3 pagesRecombinant DNA TechnologyMaika Pilley DaduboNo ratings yet
- Flashcards - Topic 2.6-2.8 Proteins and Enzymes - Edexcel IAL Biology A-LevelDocument73 pagesFlashcards - Topic 2.6-2.8 Proteins and Enzymes - Edexcel IAL Biology A-LevelJeet AminNo ratings yet
- DNA ReplicationDocument28 pagesDNA ReplicationAsim Bin Arshad 58-FBAS/BSBIO/S19100% (1)
- Cell Cycle and Cell Growth Control-2011 TextDocument68 pagesCell Cycle and Cell Growth Control-2011 TextShang-JuiTsaiNo ratings yet
- Role of DNA in Paternity TestingDocument11 pagesRole of DNA in Paternity TestingcocochilNo ratings yet
- How Cancer Cells Remodel Lipid Metabolism: Strategies Targeting Transcription FactorsDocument19 pagesHow Cancer Cells Remodel Lipid Metabolism: Strategies Targeting Transcription FactorsHossam El-basiounyNo ratings yet
- Chapter 9 Cell Communication Part 1Document40 pagesChapter 9 Cell Communication Part 1MARK BRIAN FLORESNo ratings yet
- BIOLOGY - As As-Level Biology (New Spec) CheatsheetDocument8 pagesBIOLOGY - As As-Level Biology (New Spec) CheatsheetMaie20No ratings yet
- PROTOCOLDocument4 pagesPROTOCOLfama18No ratings yet
- Gel ElectrophoresisDocument32 pagesGel ElectrophoresisSarang KottangalNo ratings yet
- IPPATH Remove and IPPOOL Add in NodeBDocument15 pagesIPPATH Remove and IPPOOL Add in NodeBasusf6veNo ratings yet
- Gene Expression: Central Dogma of Protein Synthesis: Pre-AssessmentDocument4 pagesGene Expression: Central Dogma of Protein Synthesis: Pre-AssessmentBrian NguyenNo ratings yet
- Master Students Lecture 2 Transcription and TranslationDocument28 pagesMaster Students Lecture 2 Transcription and Translationha88ial88No ratings yet
- What is the Human Genome in 40 CharactersDocument13 pagesWhat is the Human Genome in 40 CharactersRahul ThakurNo ratings yet
- Transport of Lipids in The Body FluidsDocument1 pageTransport of Lipids in The Body FluidsZeedan MohammedNo ratings yet
- Native Gel Electrophoresis: Molecular Techniques and MethodsDocument9 pagesNative Gel Electrophoresis: Molecular Techniques and MethodsSamar MustafaNo ratings yet
- Understanding HemostasisDocument53 pagesUnderstanding HemostasisGirum TesfayeNo ratings yet
- Biozone, DNA ReplicationDocument1 pageBiozone, DNA ReplicationNick BaderNo ratings yet
- Regulation of GlycogenesisDocument12 pagesRegulation of GlycogenesisCitrawati Dyah Kencono WunguNo ratings yet