You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- A Review On Advanced Oxidation Processes For The Removal of Taste and Odor Compounds From Aqueous Media PDFDocument20 pagesA Review On Advanced Oxidation Processes For The Removal of Taste and Odor Compounds From Aqueous Media PDFjose e. floresNo ratings yet
- Training ManualDocument63 pagesTraining ManualMohamad Dpr100% (1)
- Application of Different Types of Bioreactors in BioprocessesDocument37 pagesApplication of Different Types of Bioreactors in BioprocessesKarenParadaNo ratings yet
- Sharma Tamanna-2015-Lipid Content InscenedesmusDocument9 pagesSharma Tamanna-2015-Lipid Content InscenedesmusKarenParadaNo ratings yet
- Piligaev A.V-2015-Isolation of ProspectivemicroalgalDocument9 pagesPiligaev A.V-2015-Isolation of ProspectivemicroalgalKarenParadaNo ratings yet
- James C - 1995-Effect of CellDocument6 pagesJames C - 1995-Effect of CellKarenParadaNo ratings yet
- Iarael Zelitch-1997-NAS SYMPOSIUMDocument6 pagesIarael Zelitch-1997-NAS SYMPOSIUMKarenParadaNo ratings yet
- Giordano Mario-2014-Nitrogen and Sulfur Assimilation PDFDocument17 pagesGiordano Mario-2014-Nitrogen and Sulfur Assimilation PDFKarenParadaNo ratings yet
- Etan Bar-1995-PigmentDocument8 pagesEtan Bar-1995-PigmentKarenParadaNo ratings yet
- E. Gunerken-2015-Cell Disruption For MicroalgaeDocument18 pagesE. Gunerken-2015-Cell Disruption For MicroalgaeKarenParadaNo ratings yet
- Mcat Aamc Practice FLDocument7 pagesMcat Aamc Practice FLDSNo ratings yet
- A Report On The Carbon CycleDocument4 pagesA Report On The Carbon CycleJL V. AdrianoNo ratings yet
- D4209Document2 pagesD4209uocmogiandi_aNo ratings yet
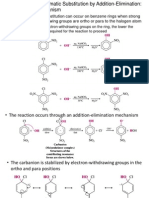
- Nucleophilic Aromatic SubstitutionDocument18 pagesNucleophilic Aromatic SubstitutionJolaine ValloNo ratings yet
- Synthetic HydrogelsDocument23 pagesSynthetic HydrogelsMohanad El-HarbawiNo ratings yet
- SASOL SURFACTANTS Product RangeDocument40 pagesSASOL SURFACTANTS Product RangeAlex100% (1)
- Exam 3 302-SolutionsDocument9 pagesExam 3 302-Solutionshuyentran1212No ratings yet
- Arkema POF Brochure 2019Document6 pagesArkema POF Brochure 2019julioNo ratings yet
- Solsperse6 2 08Document2 pagesSolsperse6 2 08Victor LopezNo ratings yet
- Hanwha Total PP HJ700Document2 pagesHanwha Total PP HJ700Nguyễn Hồng NamNo ratings yet
- Us 6645428Document20 pagesUs 6645428JohnNo ratings yet
- Understandings, Applications and Skills (This Is What You Maybe Assessed On)Document14 pagesUnderstandings, Applications and Skills (This Is What You Maybe Assessed On)Big CinemaNo ratings yet
- Energetics Question DatabaseDocument23 pagesEnergetics Question DatabaseKamrul Alam MasumNo ratings yet
- Neher 1936Document5 pagesNeher 1936M. IDRISNo ratings yet
- Chemistry SeminarDocument12 pagesChemistry SeminarBhaveshNo ratings yet
- Effect of Sugarcane Pressmud Biocompost On Dry Matter Yield and Nutrient Uptake in MaizeDocument6 pagesEffect of Sugarcane Pressmud Biocompost On Dry Matter Yield and Nutrient Uptake in MaizeNeneng May PutriNo ratings yet
- Pro Catalyst - Material Safety Data SheetDocument3 pagesPro Catalyst - Material Safety Data SheetPanya PurahongNo ratings yet
- Kendriya Vidyalaya Gachibowli, Gpra Campus, Hyd-32: Sample Paper Test 05 For Term-2 Exam (2021-22)Document12 pagesKendriya Vidyalaya Gachibowli, Gpra Campus, Hyd-32: Sample Paper Test 05 For Term-2 Exam (2021-22)Ranjeet kumar PanditNo ratings yet
- RS - Volume 9 - Issue 4 - Pages 378-391Document14 pagesRS - Volume 9 - Issue 4 - Pages 378-391samirNo ratings yet
- Molecular Biology Fourth Edition Chapt01 LectureDocument18 pagesMolecular Biology Fourth Edition Chapt01 LecturePandit JiNo ratings yet
- HydrocarbonDocument29 pagesHydrocarbondhawang40No ratings yet
- Palm Based Oleochemicals-CosmeticsDocument34 pagesPalm Based Oleochemicals-CosmeticsSharifah Zulaikha BenYahyaNo ratings yet
- Acid Rain - PP - Watermark PDFDocument15 pagesAcid Rain - PP - Watermark PDFAbhishek GuptaNo ratings yet
- Group For: A Contribution Method Second Virial CoefficientsDocument9 pagesGroup For: A Contribution Method Second Virial CoefficientsSandraColoradoNo ratings yet
- Saxena 2017Document13 pagesSaxena 2017Bimmo Dwi HartonoNo ratings yet
- Chitooligosaccharides Prevention and Control of Diseases (Se-Kwon Kim (Ed) ) PDFDocument352 pagesChitooligosaccharides Prevention and Control of Diseases (Se-Kwon Kim (Ed) ) PDFGustavo RuizNo ratings yet
- Regioselective Hydrochlorination: An Experiment For The Undergraduate LaboratoryDocument2 pagesRegioselective Hydrochlorination: An Experiment For The Undergraduate LaboratoryNgọcKhánhÔngNo ratings yet
- 15Document5 pages15gakocherNo ratings yet