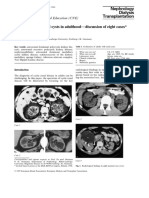
You might also like
- Tissue Engineering and Regeneration in Dentistry: Current StrategiesFrom EverandTissue Engineering and Regeneration in Dentistry: Current StrategiesRachel J. WaddingtonNo ratings yet
- Bqab 136Document12 pagesBqab 136Tatta CamposNo ratings yet
- Hereditary Podocytopathies in Adults: The Next Generation: The Kidney in Genetic and Rare Diseases: ReviewDocument7 pagesHereditary Podocytopathies in Adults: The Next Generation: The Kidney in Genetic and Rare Diseases: ReviewIgnacio TabuadaNo ratings yet
- Regan2013 Notch Signaling and Bone RemodelingDocument4 pagesRegan2013 Notch Signaling and Bone Remodelinghennysusanto18No ratings yet
- 12 Boute 00 NatGenetDocument6 pages12 Boute 00 NatGenetDorin Cristian AntalNo ratings yet
- Genetics May Hold Key to Understanding Idiopathic Nephrotic SyndromeDocument9 pagesGenetics May Hold Key to Understanding Idiopathic Nephrotic SyndromeHj herniatiNo ratings yet
- FA_Mitochondrial and NAD+ metabolism predict recovery from acute kidney injury in a diverse mouse populationDocument21 pagesFA_Mitochondrial and NAD+ metabolism predict recovery from acute kidney injury in a diverse mouse populationCher IshNo ratings yet
- B-Catetin (01-09)Document9 pagesB-Catetin (01-09)yayu latifahNo ratings yet
- B CatetinDocument20 pagesB Catetinyayu latifahNo ratings yet
- Polycystic Kidney Disease Oh2020Document10 pagesPolycystic Kidney Disease Oh2020LinaNo ratings yet
- Genes 13 00165 With CoverDocument12 pagesGenes 13 00165 With CoverMaria Garcia FernandezNo ratings yet
- A Mutation in Transcription Factor MAFB Causes Focal Segmental Glomerulosclerosis With Duane Retraction SyndromeDocument12 pagesA Mutation in Transcription Factor MAFB Causes Focal Segmental Glomerulosclerosis With Duane Retraction SyndromeCalvin Tanuwijaya Stick BolaNo ratings yet
- Ragvin PNAS10Document6 pagesRagvin PNAS10Fernando CasaresNo ratings yet
- Podocyte Biology and The EmergingDocument8 pagesPodocyte Biology and The Emerging_siegfried_No ratings yet
- Blood-Brain Barrier Pericyte Importance in Malignant GliomasDocument10 pagesBlood-Brain Barrier Pericyte Importance in Malignant GliomasMaría P SNo ratings yet
- Nephrotic Syndrome: Symposium On Pediatric NephrologyDocument11 pagesNephrotic Syndrome: Symposium On Pediatric NephrologyKumar Gavali SuryanarayanaNo ratings yet
- 2022 Janas NazeDocument10 pages2022 Janas NazeazizhamoudNo ratings yet
- Polycystic Kidney - Eight CasesDocument11 pagesPolycystic Kidney - Eight CasesKAINAT ANJUMNo ratings yet
- Frontotemporal Dementia NeuropathologyDocument14 pagesFrontotemporal Dementia Neuropathologyppico1963No ratings yet
- Animal Models of Nephrotic SyndromeDocument11 pagesAnimal Models of Nephrotic Syndromegalihrahman51No ratings yet
- Treatment of Steroid-Resistant Nephrotic Syndrome in The Genomic EraDocument15 pagesTreatment of Steroid-Resistant Nephrotic Syndrome in The Genomic EraBeatri AyuzaNo ratings yet
- Microscopy Res Technique - 2001 - Southwood - Molecular pathways of oligodendrocyte apoptosis revealed by mutations inDocument9 pagesMicroscopy Res Technique - 2001 - Southwood - Molecular pathways of oligodendrocyte apoptosis revealed by mutations inAnglia LopesNo ratings yet
- Kidney Involvement in Systemic Sclerosis JUL 2022Document15 pagesKidney Involvement in Systemic Sclerosis JUL 2022Diego Ignacio González MárquezNo ratings yet
- Role of Genetics in Orthodontics - Simplified ConceptDocument5 pagesRole of Genetics in Orthodontics - Simplified ConceptInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Apnea Del Sueñoi y Vit DDocument6 pagesApnea Del Sueñoi y Vit Dyanet guzman aybarNo ratings yet
- Neurodegenerative Diseases Multifactorial ConformationalDocument9 pagesNeurodegenerative Diseases Multifactorial ConformationalLala FemNo ratings yet
- Srep 45623Document14 pagesSrep 45623javi222222No ratings yet
- Bergmann 2018Document24 pagesBergmann 2018Jack JackNo ratings yet
- Nefrotik SyndDocument13 pagesNefrotik SyndJorianditha RamadhanNo ratings yet
- α-synuclein pathogenesis in hiPSC models of Parkinson's diseaseDocument10 pagesα-synuclein pathogenesis in hiPSC models of Parkinson's diseasecychiu1215No ratings yet
- DJ-1 Neurodegeneration Review 2021Document29 pagesDJ-1 Neurodegeneration Review 2021Krisztina TothNo ratings yet
- CurcumolDocument18 pagesCurcumolLeonardo SamuelNo ratings yet
- Cold Spring Harb Mol Case Stud-2022-Tirado-Class-a006212Document14 pagesCold Spring Harb Mol Case Stud-2022-Tirado-Class-a006212jernsssNo ratings yet
- Noonan Syndrome: Clinical Aspects and Molecular PathogenesisDocument25 pagesNoonan Syndrome: Clinical Aspects and Molecular PathogenesisFlavia ChNo ratings yet
- 2016 NGS predictive of FIBROSISDocument12 pages2016 NGS predictive of FIBROSISmaomaochongNo ratings yet
- FA_Markers of renal fibrosis- How do they correlate with podocyte damage in glomerular diseasesDocument21 pagesFA_Markers of renal fibrosis- How do they correlate with podocyte damage in glomerular diseasesCher IshNo ratings yet
- Seminar: EpidemiologyDocument15 pagesSeminar: EpidemiologyHaowen WuNo ratings yet
- Butlertjaden 2013Document15 pagesButlertjaden 2013Raluca MosoiaNo ratings yet
- Experimental Eye Research: Sarah X. Zhang, Emily Sanders, Steven J. Fliesler, Joshua J. WangDocument11 pagesExperimental Eye Research: Sarah X. Zhang, Emily Sanders, Steven J. Fliesler, Joshua J. WangJohnny GoteNo ratings yet
- Insights Into The Molecular Regulatory Network of Pathomechanisms in OsteochondromaDocument8 pagesInsights Into The Molecular Regulatory Network of Pathomechanisms in OsteochondromaEga AihenaNo ratings yet
- Semiquantitative Reverse Transcription-Polymerase Chain Reaction Analysis of Syndecan-1 and - 4 Messages in Cartilage and Cultured Chondrocytes From Osteoarthritic JointsDocument10 pagesSemiquantitative Reverse Transcription-Polymerase Chain Reaction Analysis of Syndecan-1 and - 4 Messages in Cartilage and Cultured Chondrocytes From Osteoarthritic Jointsinvestbiz optionstarNo ratings yet
- Evidence and Perspectives of Cell Senescence in Neurodegenerative DiseasesDocument11 pagesEvidence and Perspectives of Cell Senescence in Neurodegenerative DiseasesKarel GuevaraNo ratings yet
- Laminopatias PDFDocument16 pagesLaminopatias PDFSamir Fernandez RamosNo ratings yet
- 131102.2 20200108132649 CoveredDocument18 pages131102.2 20200108132649 Covered6pb4rwpk45No ratings yet
- BSR 2022 1622Document12 pagesBSR 2022 1622Maja VeselinovicNo ratings yet
- Gefs Etiologia Agatti Conh 12Document8 pagesGefs Etiologia Agatti Conh 12Leoncio SerranoNo ratings yet
- Genetic Blueprint Cardiac DevelopmentDocument6 pagesGenetic Blueprint Cardiac DevelopmentProf. David Rodriguez MartinezNo ratings yet
- Oxidative Stress-Induced Circkif18A Downregulation Impairs Mcm7-Mediated Anti-Senescence in Intervertebral Disc DegenerationDocument13 pagesOxidative Stress-Induced Circkif18A Downregulation Impairs Mcm7-Mediated Anti-Senescence in Intervertebral Disc DegenerationValen EstevezNo ratings yet
- Novel Rare Mutations in Planar Cell Polarity Gene PRICKLE1 Associated with Human Neural Tube DefectsDocument10 pagesNovel Rare Mutations in Planar Cell Polarity Gene PRICKLE1 Associated with Human Neural Tube DefectsamraovcinaNo ratings yet
- Pathologic Classification of Focal Segmental GlomerulosclerosisDocument18 pagesPathologic Classification of Focal Segmental GlomerulosclerosisRhod LadiaNo ratings yet
- Adpkd - Patho PaperDocument5 pagesAdpkd - Patho Paperapi-347957158No ratings yet
- X-Linked Juvenile Retinoschisis: Clinical Diagnosis, Genetic Analysis, and Molecular MechanismsDocument38 pagesX-Linked Juvenile Retinoschisis: Clinical Diagnosis, Genetic Analysis, and Molecular MechanismsraniNo ratings yet
- Deciphering Neurodegenerative Diseases Using Long-Read SequencingDocument12 pagesDeciphering Neurodegenerative Diseases Using Long-Read SequencingShavana RajkumarNo ratings yet
- CADAE1405CDocument12 pagesCADAE1405CMuhammad RiduanNo ratings yet
- Hereditary Podocytopathies in Adults: The Next GenerationDocument17 pagesHereditary Podocytopathies in Adults: The Next Generationpkbljr sulutNo ratings yet
- Sindrome NefroticoDocument8 pagesSindrome NefroticoRudy Espejo del ArcaNo ratings yet
- Author: Section Editor: Deputy Editor:: Pathogenesis of Systemic Sclerosis (Scleroderma)Document24 pagesAuthor: Section Editor: Deputy Editor:: Pathogenesis of Systemic Sclerosis (Scleroderma)zakiNo ratings yet
- JURNALDocument10 pagesJURNALLuses Shantia HaryantoNo ratings yet
- Microglial Phagocytosis of Neurons in Neurodegeneration, and Its RegulationDocument19 pagesMicroglial Phagocytosis of Neurons in Neurodegeneration, and Its RegulationChristina MountakiNo ratings yet
- The Podocyte's Response to Injury: Role in Proteinuria and GlomerulosclerosisDocument17 pagesThe Podocyte's Response to Injury: Role in Proteinuria and Glomerulosclerosisgalihrahman51No ratings yet
- Ku'i Ka Pahu: Regional Director's Message by RD Sharon Ishida, 2020-2021Document6 pagesKu'i Ka Pahu: Regional Director's Message by RD Sharon Ishida, 2020-2021Ignacio TabuadaNo ratings yet
- Advance Google Search Techniques Using "Google DorksDocument13 pagesAdvance Google Search Techniques Using "Google DorksASLAM-O- ALAKUMNo ratings yet
- The Bike Shop Had Special Bike Racks Out Front, Which Were Shaped Like Bicycle Wheels.Document204 pagesThe Bike Shop Had Special Bike Racks Out Front, Which Were Shaped Like Bicycle Wheels.Ignacio TabuadaNo ratings yet
- Feasibility Study On Production and Sustainability of Poly Propylene Fiber Reinforced Concrete Ties Based On A Value Engineering SurveyDocument8 pagesFeasibility Study On Production and Sustainability of Poly Propylene Fiber Reinforced Concrete Ties Based On A Value Engineering SurveyIgnacio TabuadaNo ratings yet
- She Knew She Should Be Grateful But Still Felt Sad.Document6 pagesShe Knew She Should Be Grateful But Still Felt Sad.Ignacio TabuadaNo ratings yet
- Central Y Service Club Newsletter Highlights Community ServiceDocument2 pagesCentral Y Service Club Newsletter Highlights Community ServiceIgnacio TabuadaNo ratings yet
- Dissertation Zur Erlangung Des DoktorgradesDocument164 pagesDissertation Zur Erlangung Des DoktorgradesIgnacio TabuadaNo ratings yet
- Recombinant Human Resistin ProteinDocument2 pagesRecombinant Human Resistin ProteinIgnacio TabuadaNo ratings yet
- Home Keys2Document1 pageHome Keys2Ignacio TabuadaNo ratings yet
- Caap 92 4 0Document105 pagesCaap 92 4 0Ignacio TabuadaNo ratings yet
- Central 1303Document2 pagesCentral 1303Ignacio TabuadaNo ratings yet
- Bingocards Free SampleDocument4 pagesBingocards Free SampleIgnacio TabuadaNo ratings yet
- Suspicious Board Meeting Notice Raises DoubtDocument6 pagesSuspicious Board Meeting Notice Raises DoubtIgnacio TabuadaNo ratings yet
- PRDF MST720C-LFDocument127 pagesPRDF MST720C-LFIgnacio Tabuada0% (1)
- Msa SS01517Document3 pagesMsa SS01517Ignacio TabuadaNo ratings yet
- TB Lecture41 ETABS Load CombinationsDocument12 pagesTB Lecture41 ETABS Load CombinationsIgnacio TabuadaNo ratings yet
- Msa SS01517Document3 pagesMsa SS01517Ignacio TabuadaNo ratings yet
- Steel Structural Design Using Nonlinear Inelastic Analysis: Viet-Hung Truong and Seung-Eock KimDocument9 pagesSteel Structural Design Using Nonlinear Inelastic Analysis: Viet-Hung Truong and Seung-Eock KimIgnacio TabuadaNo ratings yet
- 1950 04 07 HistorySNDocument76 pages1950 04 07 HistorySNIgnacio TabuadaNo ratings yet
- ICAO Aerodrome Certification RequirementsDocument24 pagesICAO Aerodrome Certification RequirementsIgnacio TabuadaNo ratings yet
- NCLB-ADC WS-Training-PPT5-Regulator Certification ExperienceDocument41 pagesNCLB-ADC WS-Training-PPT5-Regulator Certification ExperienceIgnacio TabuadaNo ratings yet
- Airlines CodesDocument11 pagesAirlines CodesJennifer Zambrano100% (1)
- Master PlanningDocument3 pagesMaster PlanningIgnacio TabuadaNo ratings yet
- 1.4 ICAO AMPTF - AIRSIDE DEVELOPMENT (MPlarre) PDFDocument20 pages1.4 ICAO AMPTF - AIRSIDE DEVELOPMENT (MPlarre) PDFVasaNo ratings yet
- V Wicher PDFDocument1 pageV Wicher PDFIgnacio TabuadaNo ratings yet
- Designing For Lateral-Torsional Stability in Wood MembersDocument10 pagesDesigning For Lateral-Torsional Stability in Wood MembersIgnacio TabuadaNo ratings yet
- ICAO Current Work On Aerodrome Planning PDFDocument54 pagesICAO Current Work On Aerodrome Planning PDFUmar FarooqNo ratings yet
- Nonlinear Inelastic Analysis of Steel-Concrete Composite Beam-Columns Using The Stability FunctionsDocument23 pagesNonlinear Inelastic Analysis of Steel-Concrete Composite Beam-Columns Using The Stability FunctionsIgnacio TabuadaNo ratings yet
- Addendum No. 1 - 6Document3 pagesAddendum No. 1 - 6Ignacio TabuadaNo ratings yet
- Evolution and State of The Art of Column Stability CriteriaDocument8 pagesEvolution and State of The Art of Column Stability CriteriaIgnacio TabuadaNo ratings yet
- En 10025 PDFDocument35 pagesEn 10025 PDFAbdelali88% (8)
- 9th Science Mcqs PTB PDFDocument25 pages9th Science Mcqs PTB PDFUmar JuttNo ratings yet
- Marine Environmental ResearchDocument11 pagesMarine Environmental ResearchPierre Otoniel Carmen MunaycoNo ratings yet
- Below Is A List of Offices For India - PWCDocument10 pagesBelow Is A List of Offices For India - PWCAnonymous xMYE0TiNBcNo ratings yet
- Soal Mid Ganjil 14Document4 pagesSoal Mid Ganjil 14Anonymous a2C6YgevfNo ratings yet
- Organizational Behavior: Eighteenth EditionDocument37 pagesOrganizational Behavior: Eighteenth EditionPrashant Kumar100% (1)
- Oracle Analytics Cloud 2018 Solution Engineer Specialist AssessmentDocument4 pagesOracle Analytics Cloud 2018 Solution Engineer Specialist AssessmentRamón MedinaNo ratings yet
- DocxDocument5 pagesDocxFelicya Angel Ivy LynnNo ratings yet
- Shahrzadgh InquiryassignmentDocument5 pagesShahrzadgh Inquiryassignmentapi-274076411No ratings yet
- 11g SQL Fundamentals I Student Guide - Vol IIDocument283 pages11g SQL Fundamentals I Student Guide - Vol IIIjazKhanNo ratings yet
- CaffeineDocument2 pagesCaffeineSaini Malkeet100% (1)
- Sindhi 2020Document1 pageSindhi 2020Engr Javed AkhtarNo ratings yet
- IN THE COURT OF - SUIT/APPEAL No. - JURISDICTION OF 2017Document1 pageIN THE COURT OF - SUIT/APPEAL No. - JURISDICTION OF 2017Adv Pooja AroraNo ratings yet
- Business Studies Project: Made By: Rahil JainDocument29 pagesBusiness Studies Project: Made By: Rahil JainChirag KothariNo ratings yet
- Refunds Maceda Law and PD957Document2 pagesRefunds Maceda Law and PD957QUINTO CRISTINA MAENo ratings yet
- Importance of family history and learning about ancestorsDocument2 pagesImportance of family history and learning about ancestorsKei ArceñoNo ratings yet
- (English (Auto-Generated) ) Reviving The Labor Movement - Margaret Levi - TEDxSeattle (DownSub - Com)Document14 pages(English (Auto-Generated) ) Reviving The Labor Movement - Margaret Levi - TEDxSeattle (DownSub - Com)Surat PEHKTNo ratings yet
- Biology and Aquaculture of Tilapia-Routledge (2022)Document324 pagesBiology and Aquaculture of Tilapia-Routledge (2022)Pablo Antonio Pintos TeránNo ratings yet
- GasGas TXT Racing 2014 Owner's ManualDocument60 pagesGasGas TXT Racing 2014 Owner's Manualstone chouNo ratings yet
- Late 18th and Early 19thDocument82 pagesLate 18th and Early 19thGrace CordelinNo ratings yet
- PDF Penstock Manual DLDocument160 pagesPDF Penstock Manual DLWilmer Fernando DuarteNo ratings yet
- Lcasean PaperDocument6 pagesLcasean Paperkean ebeoNo ratings yet
- Guide to Preparing STEM Fellowship ApplicationsDocument14 pagesGuide to Preparing STEM Fellowship ApplicationsNurrahmiNo ratings yet
- Distribution and Production of Rice in IndiaDocument6 pagesDistribution and Production of Rice in IndiaZehan SheikhNo ratings yet
- EM Complete NotesDocument191 pagesEM Complete NoteswsbwaqhpwpvcwuhajkNo ratings yet
- Call For Applications To JASTIP-Net 2021 Guidelines For ApplicantsDocument6 pagesCall For Applications To JASTIP-Net 2021 Guidelines For ApplicantsNadya IrsalinaNo ratings yet
- SL - No Name of The Students University Roll NumberDocument58 pagesSL - No Name of The Students University Roll NumberSai AmbatiNo ratings yet
- The Concept of Social Capital: A Critical Review: Sophie Ponthieux ( )Document23 pagesThe Concept of Social Capital: A Critical Review: Sophie Ponthieux ( )Ali Muhtarom LilikNo ratings yet
- Dell M1000e Install Admin TroubleshootingDocument78 pagesDell M1000e Install Admin Troubleshootingsts100No ratings yet
- Gabriel's Oboe String PartsDocument5 pagesGabriel's Oboe String Partshekerer50% (2)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationFrom EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationRating: 4 out of 5 stars4/5 (1)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- Fearfully and Wonderfully: The Marvel of Bearing God's ImageFrom EverandFearfully and Wonderfully: The Marvel of Bearing God's ImageRating: 5 out of 5 stars5/5 (40)
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceFrom EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceRating: 4 out of 5 stars4/5 (632)