You might also like
- NROSCI 1012 - Lecture 23Document3 pagesNROSCI 1012 - Lecture 23HonzaNo ratings yet
- TMP 9 DA2Document11 pagesTMP 9 DA2FrontiersNo ratings yet
- SynapseDocument48 pagesSynapseSaurabh KrishnaNo ratings yet
- NROSCI 1012 - Lecture 24Document4 pagesNROSCI 1012 - Lecture 24HonzaNo ratings yet
- Synaptic Depression: Many Different Possible MechanismsDocument24 pagesSynaptic Depression: Many Different Possible MechanismsThe Panda EntertainerNo ratings yet
- Neurobiology 4 - A - Synaptic PlasticityDocument26 pagesNeurobiology 4 - A - Synaptic PlasticitymolinakevinalejandroNo ratings yet
- 12 Synaptic Plasticity STDPDocument3 pages12 Synaptic Plasticity STDPCesia ConstanzaNo ratings yet
- Calcium Control of Neurotransmitter ReleaseDocument15 pagesCalcium Control of Neurotransmitter ReleaseNisa Yulianti SuprahmanNo ratings yet
- Calcium Control of Neurotransmitter Release PDFDocument15 pagesCalcium Control of Neurotransmitter Release PDFNisa Yulianti SuprahmanNo ratings yet
- Cellular Mechanisms of Cerebellar LTD: Hervé Daniel, Carole Levenes and Francis CrépelDocument7 pagesCellular Mechanisms of Cerebellar LTD: Hervé Daniel, Carole Levenes and Francis Crépelrocambolescas perthNo ratings yet
- 01 SynapseDocument15 pages01 SynapseT. ManopriyaNo ratings yet
- LTP1 ( 2 Hours, 1 TBS) Depends On Modifications of Postsynaptic Proteins (MainlyDocument12 pagesLTP1 ( 2 Hours, 1 TBS) Depends On Modifications of Postsynaptic Proteins (MainlyThe Panda EntertainerNo ratings yet
- Lecture 8ADocument17 pagesLecture 8AAbdullah EwaisNo ratings yet
- 28 Optical - Techniques PDFDocument28 pages28 Optical - Techniques PDFThe Panda EntertainerNo ratings yet
- Jurnal b6Document24 pagesJurnal b6Fatimah Hartina FaradillahNo ratings yet
- SynapsesDocument6 pagesSynapsesJeff ParkNo ratings yet
- Goldschmidt Clermont2015Document8 pagesGoldschmidt Clermont2015marcela gomezNo ratings yet
- IndexDocument123 pagesIndexbszool006No ratings yet
- Synapses and Synaptic TransmissionDocument19 pagesSynapses and Synaptic TransmissionSabinaNo ratings yet
- Zheng 2006 NatNeuro Bursting Intrinsic Retina PDFDocument9 pagesZheng 2006 NatNeuro Bursting Intrinsic Retina PDFSimón Salgado MuñozNo ratings yet
- Synaptic TransmissionDocument17 pagesSynaptic TransmissionbnmjgcNo ratings yet
- Neurobiology Lectures 5-6Document58 pagesNeurobiology Lectures 5-6Saajid AmraNo ratings yet
- Synaptic Transmission: Transmission at The Synapses May BeDocument13 pagesSynaptic Transmission: Transmission at The Synapses May Beramadan100% (3)
- Neurobiology Lectures 5-6Document58 pagesNeurobiology Lectures 5-6Saajid AmraNo ratings yet
- The Synapse: DefintionDocument55 pagesThe Synapse: DefintionQairul Azman100% (1)
- M - PH: The Relationship Between Length and Active Force of A Skeletal MuscleDocument11 pagesM - PH: The Relationship Between Length and Active Force of A Skeletal MuscleFlowerNo ratings yet
- Carrier-Envelope Phase Control of Few-Cycle Parametric Chirped-Pulse AmplifierDocument5 pagesCarrier-Envelope Phase Control of Few-Cycle Parametric Chirped-Pulse AmplifierDu RoyNo ratings yet
- 20 Synaptic Transmission - Vel - Ca - Nano Vs Microdomains PDFDocument23 pages20 Synaptic Transmission - Vel - Ca - Nano Vs Microdomains PDFThe Panda EntertainerNo ratings yet
- SynapsesDocument46 pagesSynapseshan abuur ahmedNo ratings yet
- 11 Synaptic Plasticity LTP LTDDocument4 pages11 Synaptic Plasticity LTP LTDCesia ConstanzaNo ratings yet
- Local AnestheticsDocument30 pagesLocal AnestheticsmaurinmarceoiaNo ratings yet
- Clayton 1997 0185Document3 pagesClayton 1997 0185Particle Beam Physics LabNo ratings yet
- Pharmacology of Central Nervous SystemDocument20 pagesPharmacology of Central Nervous SystemAngelic khan100% (1)
- Section 2: Signal Transmission Between The NeuronsDocument123 pagesSection 2: Signal Transmission Between The NeuronsAna Crigan VlasNo ratings yet
- ANTI ARRHTYMIC DRUG IncompleteDocument9 pagesANTI ARRHTYMIC DRUG IncompleteJustine CastilloNo ratings yet
- Lecture 2Document16 pagesLecture 2Razvan NegritasNo ratings yet
- Chapter #55 - Physiology and Pharmacology of The Renal Pelvis and UreterDocument6 pagesChapter #55 - Physiology and Pharmacology of The Renal Pelvis and UreterMohammadSAL-RawashdehNo ratings yet
- Repetitve Nerve Stimulation (RNS) : By: Syed Irshad Murtaza Neurophysiology Dept AKUH Karachi Date:12-06-2013Document33 pagesRepetitve Nerve Stimulation (RNS) : By: Syed Irshad Murtaza Neurophysiology Dept AKUH Karachi Date:12-06-2013Raghu NadhNo ratings yet
- Dr. Prayogo NBS 2019Document25 pagesDr. Prayogo NBS 2019Arrafi GanNo ratings yet
- Synaptic Amplification in Motoneurons Computational and Mechanistic ImplicationsDocument3 pagesSynaptic Amplification in Motoneurons Computational and Mechanistic ImplicationsdagushNo ratings yet
- 06 SynapseDocument22 pages06 SynapseJaydave PatelNo ratings yet
- The Journal of Neuroscience, July 1992, 12 (7) : 2516-2522Document7 pagesThe Journal of Neuroscience, July 1992, 12 (7) : 2516-2522Carrie PartchNo ratings yet
- Synapses and Synaptic Transmission: Dr. Eman El Eter Physiology Dep. College of Medicine KSUDocument43 pagesSynapses and Synaptic Transmission: Dr. Eman El Eter Physiology Dep. College of Medicine KSUraanja2No ratings yet
- CH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewDocument4 pagesCH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewoguzNo ratings yet
- Week 10 Comprehensive ReviewDocument4 pagesWeek 10 Comprehensive Reviewandreil12345No ratings yet
- Gennady Shvets Et Al - Excitation of Accelerating Plasma Waves by Counter-Propagating Laser BeamsDocument10 pagesGennady Shvets Et Al - Excitation of Accelerating Plasma Waves by Counter-Propagating Laser BeamsDublin000No ratings yet
- Cheat ECGDocument3 pagesCheat ECGNeil Emman BotardoNo ratings yet
- Neuromuscular Junction Physiology & Blocking Agents: Prof V K Bhatia Dept of Anaesthesiology KgmuDocument39 pagesNeuromuscular Junction Physiology & Blocking Agents: Prof V K Bhatia Dept of Anaesthesiology KgmuAthira S MadhuNo ratings yet
- 1 s2.0 S0960982205006184 MainDocument7 pages1 s2.0 S0960982205006184 MainPhysics WANGNo ratings yet
- A 23 GHZ Fast-Locking PLL Using Phase Error CompensatorDocument5 pagesA 23 GHZ Fast-Locking PLL Using Phase Error Compensatornayakadarsh2000No ratings yet
- SynapseDocument3 pagesSynapsebszool006No ratings yet
- Article: Release-Dependent Variations in Synaptic Latency: A Putative Code For Short-And Long-Term Synaptic DynamicsDocument13 pagesArticle: Release-Dependent Variations in Synaptic Latency: A Putative Code For Short-And Long-Term Synaptic DynamicsDi Fon Panse'wNo ratings yet
- Wolf ParkinsonDocument4 pagesWolf ParkinsonAlejandro Bañol EscobarNo ratings yet
- Molecular Pain: Short-Term Synaptic Plasticity in The Nociceptive Thalamic-Anterior Cingulate PathwayDocument20 pagesMolecular Pain: Short-Term Synaptic Plasticity in The Nociceptive Thalamic-Anterior Cingulate PathwayYixin TongNo ratings yet
- Nihms 181336Document6 pagesNihms 181336Giulia AndreeaNo ratings yet
- MCB 80 - Section Week 3 Worksheet TF: Alix Lacoste Quantal AnalysisDocument2 pagesMCB 80 - Section Week 3 Worksheet TF: Alix Lacoste Quantal AnalysisCARDENAS MARIA ROSA YOLITZYNo ratings yet
- Suk 2001 0431Document4 pagesSuk 2001 0431Particle Beam Physics LabNo ratings yet
- Introduction To Peripheral Nervous System PharmacologyDocument30 pagesIntroduction To Peripheral Nervous System PharmacologyYolanda SimamoraNo ratings yet
- PET in Neurological DisordersDocument12 pagesPET in Neurological Disordersalfred benedict bayanNo ratings yet
- UNIEURO InvestorPresentation Jan2020 3 2 PDFDocument47 pagesUNIEURO InvestorPresentation Jan2020 3 2 PDFThe Panda EntertainerNo ratings yet
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Unieuro FY2018Document411 pagesUnieuro FY2018The Panda EntertainerNo ratings yet
- UNIEURO InvestorPresentation Jan2020 3 2 PDFDocument47 pagesUNIEURO InvestorPresentation Jan2020 3 2 PDFThe Panda EntertainerNo ratings yet
- The Shift in Global Crude Oil Market Structure: A Model-Based Analysis of The Period 2013-2017Document39 pagesThe Shift in Global Crude Oil Market Structure: A Model-Based Analysis of The Period 2013-2017The Panda EntertainerNo ratings yet
- Unieuro InvestorPresentation May17 - 1Document54 pagesUnieuro InvestorPresentation May17 - 1The Panda EntertainerNo ratings yet
- Company Profile - 2 PDFDocument1 pageCompany Profile - 2 PDFThe Panda EntertainerNo ratings yet
- 26 Learning - Memory2 - LTP1 PDFDocument13 pages26 Learning - Memory2 - LTP1 PDFThe Panda EntertainerNo ratings yet
- Interim Directors' Report: As at 30 November 2019Document31 pagesInterim Directors' Report: As at 30 November 2019The Panda EntertainerNo ratings yet
- Unieuro HalfYearFinancialReport2019Document97 pagesUnieuro HalfYearFinancialReport2019The Panda EntertainerNo ratings yet
- UNIEURO FY1718ResultsnnDocument37 pagesUNIEURO FY1718ResultsnnThe Panda EntertainerNo ratings yet
- 28 Optical - Techniques PDFDocument28 pages28 Optical - Techniques PDFThe Panda EntertainerNo ratings yet
- UNIEURO AnnualFinancialReport 2018 PDFDocument410 pagesUNIEURO AnnualFinancialReport 2018 PDFThe Panda EntertainerNo ratings yet
- Neurobiology Assignment PDFDocument2 pagesNeurobiology Assignment PDFThe Panda EntertainerNo ratings yet
- Mechanisms of Short-Term Synaptic PlasticityDocument13 pagesMechanisms of Short-Term Synaptic PlasticityThe Panda EntertainerNo ratings yet
- 8 Cortex4 - Associative - Motor PDFDocument20 pages8 Cortex4 - Associative - Motor PDFThe Panda EntertainerNo ratings yet
- 15 Firing Mechanisms - KCa PDFDocument11 pages15 Firing Mechanisms - KCa PDFThe Panda EntertainerNo ratings yet
- 25 Learning - Memory1 PDFDocument28 pages25 Learning - Memory1 PDFThe Panda EntertainerNo ratings yet
- 21 Synaptic Plasticity - Short - Term PDFDocument19 pages21 Synaptic Plasticity - Short - Term PDFThe Panda EntertainerNo ratings yet
- 13 Firing Mechanisms - Voltage - Clamp - Protocols2 - Neuro20 PDFDocument37 pages13 Firing Mechanisms - Voltage - Clamp - Protocols2 - Neuro20 PDFThe Panda EntertainerNo ratings yet
- 20 Synaptic Transmission - Vel - Ca - Nano Vs Microdomains PDFDocument23 pages20 Synaptic Transmission - Vel - Ca - Nano Vs Microdomains PDFThe Panda EntertainerNo ratings yet
- 14 Firing Mechanisms - LVA Channels PDFDocument20 pages14 Firing Mechanisms - LVA Channels PDFThe Panda EntertainerNo ratings yet
- 10 Basal - Ganglia PDFDocument11 pages10 Basal - Ganglia PDFThe Panda EntertainerNo ratings yet
- 7 Cortex3 PDFDocument20 pages7 Cortex3 PDFThe Panda EntertainerNo ratings yet
- 9 Cerebellum PDFDocument25 pages9 Cerebellum PDFThe Panda EntertainerNo ratings yet
- 4 Brainstem PDFDocument10 pages4 Brainstem PDFThe Panda EntertainerNo ratings yet
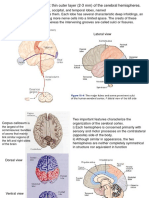
- Thin Outer Layer (2-3 MM) of The Cerebral HemispheresDocument15 pagesThin Outer Layer (2-3 MM) of The Cerebral HemispheresThe Panda EntertainerNo ratings yet
- 12 Firing Mechanisms - Voltage Clamp - Protocols - 20 PDFDocument26 pages12 Firing Mechanisms - Voltage Clamp - Protocols - 20 PDFThe Panda EntertainerNo ratings yet
- Dorsal Part of The DiencephalonDocument7 pagesDorsal Part of The DiencephalonThe Panda EntertainerNo ratings yet
- Lab 5Document12 pagesLab 5api-272723910No ratings yet
- Neurobiology 4 - A - Synaptic PlasticityDocument26 pagesNeurobiology 4 - A - Synaptic PlasticitymolinakevinalejandroNo ratings yet
- Phys - 1S04 - SynapseDocument9 pagesPhys - 1S04 - SynapseTMC PGI GENER MICKONo ratings yet
- Mechanism of Brain Changes in AddictionDocument133 pagesMechanism of Brain Changes in AddictionJared FeldmanNo ratings yet
- Articulo KANDEL - The Molecular and Systems Biology of Memory PDFDocument29 pagesArticulo KANDEL - The Molecular and Systems Biology of Memory PDFKarina NoelNo ratings yet
- Memory PhysiologyDocument30 pagesMemory PhysiologyMudassar RoomiNo ratings yet
- Memory PhysiologyDocument40 pagesMemory PhysiologyAttiqaQureshiNo ratings yet
- CNS PDFDocument412 pagesCNS PDFSami Juggy G100% (1)
- PNF PatternsDocument19 pagesPNF PatternsDany VirgilNo ratings yet
- Gladys Cheing RS3030 CNN Introduction To Neuroscience Note 2021Document73 pagesGladys Cheing RS3030 CNN Introduction To Neuroscience Note 2021Tsang AmyNo ratings yet
- Metal-Oxide Heterojunction: From Material Process To Neuromorphic ApplicationsDocument23 pagesMetal-Oxide Heterojunction: From Material Process To Neuromorphic ApplicationsjjNo ratings yet
- General PsychiatryDocument836 pagesGeneral PsychiatryGEORG100% (2)
- Synaptic Transmission at The Crayfish Superficial Flexor Muscle and Third NerveDocument14 pagesSynaptic Transmission at The Crayfish Superficial Flexor Muscle and Third Nerveapi-340131743No ratings yet
- Li Et Al 2003Document6 pagesLi Et Al 2003Adrian McDoucheNo ratings yet
- Crayfish Lab Report FinalDocument20 pagesCrayfish Lab Report Finalapi-307968207No ratings yet
- Neuromorphic Engineering: From Biological To Spike Based Hardware Nervous SystemsDocument33 pagesNeuromorphic Engineering: From Biological To Spike Based Hardware Nervous SystemsSruthy sureshNo ratings yet
- Thoughts, Consciousness and MemoryDocument28 pagesThoughts, Consciousness and MemoryJorge De VeraNo ratings yet