0% found this document useful (0 votes)
156 views12 pagesUnderstanding Flower Structure and Function
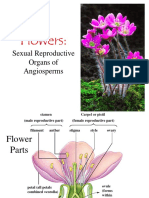
Flowers are the reproductive structures of flowering plants. They contain reproductive organs and develop from flowers to produce fruit containing seeds. Flowers come in a diverse range of shapes, sizes, and colors to attract pollinators like insects and birds. Their reproductive parts include sepals, petals, stamens that produce pollen, and a pistil with an ovary, style and stigma. Flowers are adapted for pollination through mechanisms like pollen transfer by animals or wind. After fertilization, the ovary develops into fruit housing seeds.
Uploaded by
jolinmartinCopyright
© © All Rights Reserved
We take content rights seriously. If you suspect this is your content, claim it here.
Available Formats
Download as PDF, TXT or read online on Scribd
0% found this document useful (0 votes)
156 views12 pagesUnderstanding Flower Structure and Function
Flowers are the reproductive structures of flowering plants. They contain reproductive organs and develop from flowers to produce fruit containing seeds. Flowers come in a diverse range of shapes, sizes, and colors to attract pollinators like insects and birds. Their reproductive parts include sepals, petals, stamens that produce pollen, and a pistil with an ovary, style and stigma. Flowers are adapted for pollination through mechanisms like pollen transfer by animals or wind. After fertilization, the ovary develops into fruit housing seeds.
Uploaded by
jolinmartinCopyright
© © All Rights Reserved
We take content rights seriously. If you suspect this is your content, claim it here.
Available Formats
Download as PDF, TXT or read online on Scribd