You might also like
- Evita Infinity v500 SW 2n Ifu 9052161 Ingles PDFDocument384 pagesEvita Infinity v500 SW 2n Ifu 9052161 Ingles PDFTu Papa2411No ratings yet
- Preliminary Design Review - Ad Lunam HopperDocument100 pagesPreliminary Design Review - Ad Lunam HopperAniketh DevarasettyNo ratings yet
- Fluid Mechanics Methods GuideDocument281 pagesFluid Mechanics Methods GuideJason StanleyNo ratings yet
- Camporeale 2013Document36 pagesCamporeale 2013Angel ATNo ratings yet
- 10.1007@s40808 020 00774 1Document11 pages10.1007@s40808 020 00774 1hanaeNo ratings yet
- Marani Et Al. 2013Document5 pagesMarani Et Al. 2013Jason StanleyNo ratings yet
- Restrepo++ 2015 Anthro-010Document16 pagesRestrepo++ 2015 Anthro-010Juan Sebastian Roncancio BaqueroNo ratings yet
- El Ouahli Bis STDocument12 pagesEl Ouahli Bis STEl Youbi MohammedNo ratings yet
- Bord Eyne 2017Document13 pagesBord Eyne 2017Brigita de BrillarNo ratings yet
- 1 s2.0 S0037073820301974 MainDocument11 pages1 s2.0 S0037073820301974 MainCris FischerNo ratings yet
- 10008-Article Text-56143-2-10-20211213Document9 pages10008-Article Text-56143-2-10-20211213philosophy-thoughtNo ratings yet
- Analyzing Water Quality and Eutrophication Trends in Laguna de BayDocument13 pagesAnalyzing Water Quality and Eutrophication Trends in Laguna de BayKathleen Joyce AustriaNo ratings yet
- Marine Pollution Bulletin: Bueno C., Brugnoli E., Bergamino L., Muniz P., García-Rodríguez F., Figueira RDocument7 pagesMarine Pollution Bulletin: Bueno C., Brugnoli E., Bergamino L., Muniz P., García-Rodríguez F., Figueira RThiago SilveiraNo ratings yet
- Effects Ofbottomtopographyandanthropogenicpressureon Northern Adriatic Cystoseira Spp. (Phaeophyceae, Fucales)Document7 pagesEffects Ofbottomtopographyandanthropogenicpressureon Northern Adriatic Cystoseira Spp. (Phaeophyceae, Fucales)ZvijezdaNo ratings yet
- In Uence of Hydrology Process On Wetland Landscape Pattern: A Case Study in The Yellow River DeltaDocument8 pagesIn Uence of Hydrology Process On Wetland Landscape Pattern: A Case Study in The Yellow River DeltaLaura Ovia SausaennNo ratings yet
- PRI 11 2014 LalandeDocument18 pagesPRI 11 2014 LalandeNu LeeNo ratings yet
- Pletterbauer 2014 Impact CC Fish EU RiversDocument20 pagesPletterbauer 2014 Impact CC Fish EU RiversMohammad LakNo ratings yet
- Impacts of Climate Change on Biodiversity and Ecosystem ServicesDocument8 pagesImpacts of Climate Change on Biodiversity and Ecosystem ServicesRanjeet LamaNo ratings yet
- Vortices, Lagrangian & OceanDocument17 pagesVortices, Lagrangian & OceanBryan GraczykNo ratings yet
- CliFEM - Climate Forcing and Erosion Modelling at Long-Term Sele River Research Basin (Southern Italy)Document10 pagesCliFEM - Climate Forcing and Erosion Modelling at Long-Term Sele River Research Basin (Southern Italy)Nazzareno DiodatoNo ratings yet
- Wave-Induced Velocities Inside A Model Seagrass Bed: CitationDocument16 pagesWave-Induced Velocities Inside A Model Seagrass Bed: CitationGIRISHA001No ratings yet
- Ismej 2011132 ADocument12 pagesIsmej 2011132 Aamira yaakopNo ratings yet
- Kim et al., 20bDocument14 pagesKim et al., 20bbaptistedupuNo ratings yet
- 2014-Implementing the DPSIR framework to link water quality of rivers to land use- methodological issues and preliminary field testDocument18 pages2014-Implementing the DPSIR framework to link water quality of rivers to land use- methodological issues and preliminary field testSaid RamdlanNo ratings yet
- Batalla Etal 2017 GeomorphologyDocument16 pagesBatalla Etal 2017 GeomorphologyTyP ConsultoresNo ratings yet
- ThreatsFishBiodiversity PDFDocument12 pagesThreatsFishBiodiversity PDFLucía SolerNo ratings yet
- tmpBAA5 TMPDocument10 pagestmpBAA5 TMPFrontiersNo ratings yet
- 1 s2.0 S2666916122000147 MainDocument12 pages1 s2.0 S2666916122000147 MainAbdelhak AlmNo ratings yet
- Data-driven models capture coastal aquifer salinizationDocument11 pagesData-driven models capture coastal aquifer salinizationAdi adeNo ratings yet
- Climate change impacts on Tagus estuary salinityDocument4 pagesClimate change impacts on Tagus estuary salinityWilliem HartonoNo ratings yet
- Araújo Et Al. - 2017 Inter Annual Changes in Fish Communities of A Tropical Bay BrazilDocument12 pagesAraújo Et Al. - 2017 Inter Annual Changes in Fish Communities of A Tropical Bay BrazilJuliana AbreuNo ratings yet
- Eutrophication and Restoration of Shallow LakesDocument7 pagesEutrophication and Restoration of Shallow LakesEmilia RaduNo ratings yet
- BG 19 4035 2022Document31 pagesBG 19 4035 2022MarrcoNo ratings yet
- Effect of Light and Sediment Grain Size On The Vertical Migration of Benthic DiatomsDocument8 pagesEffect of Light and Sediment Grain Size On The Vertical Migration of Benthic DiatomsCris FischerNo ratings yet
- 12 - Future of Mangrove Ecosystems To 2025 Pp. 172-187Document17 pages12 - Future of Mangrove Ecosystems To 2025 Pp. 172-187Soma GhoshNo ratings yet
- Journal of Environmental ManagementDocument14 pagesJournal of Environmental ManagementNicholas de LeónNo ratings yet
- Chapter 2 Distribution and Habitats of Aquatic InsectsDocument23 pagesChapter 2 Distribution and Habitats of Aquatic InsectsIvan GonzalezNo ratings yet
- Marine Environmental Research: T.F. Grilo, P.G. Cardoso, M.A. PardalDocument7 pagesMarine Environmental Research: T.F. Grilo, P.G. Cardoso, M.A. PardalMaria LCNo ratings yet
- An Approximate Solution To The Flow Field On Vegetated Intertidal Platforms: Applicability and LimitationsDocument22 pagesAn Approximate Solution To The Flow Field On Vegetated Intertidal Platforms: Applicability and LimitationsTomas Van OyenNo ratings yet
- Girum, T.Document12 pagesGirum, T.Arega GenetieNo ratings yet
- Impact of Biotic and Abiotic Processes On Sediment Dynamics and The Consequences To The Structure and Functioning of The Intertidal ZoneDocument14 pagesImpact of Biotic and Abiotic Processes On Sediment Dynamics and The Consequences To The Structure and Functioning of The Intertidal ZoneKha LidNo ratings yet
- Francisetal AquaticSciencesinpressDocument16 pagesFrancisetal AquaticSciencesinpressdanielaNo ratings yet
- Anot Gobbi Qualification Diego Casas 20221031Document35 pagesAnot Gobbi Qualification Diego Casas 20221031mfgobbi4515No ratings yet
- Chemical Composition of Suspended SedimeDocument16 pagesChemical Composition of Suspended SedimeLIYOUCK Patrice RolandNo ratings yet
- Govin2012 Terrigenous Input RatiosDocument23 pagesGovin2012 Terrigenous Input RatiosPaulo CettoNo ratings yet
- Factors ControllingDocument9 pagesFactors ControllingOscar BlancoNo ratings yet
- Articulo 5Document9 pagesArticulo 5Cirene Guisell Bolaño JimenezNo ratings yet
- Modeling influences of biology and physics on intertidal sediment transportDocument8 pagesModeling influences of biology and physics on intertidal sediment transportjorgepierNo ratings yet
- Impact of A Harbour Construction On The Benthic Community of TwoDocument11 pagesImpact of A Harbour Construction On The Benthic Community of TwoShanly ConeoNo ratings yet
- PDF Original 3Document12 pagesPDF Original 3MARCO ENCISO HUAMANNo ratings yet
- Taufik AmbioDocument13 pagesTaufik Ambiomobster44No ratings yet
- Van Der Heijden and Kamenos 2015Document13 pagesVan Der Heijden and Kamenos 2015Amanda CoraçãoNo ratings yet
- Benedetti 2016 Climate Change Impact On Planktonic Production in The Mediterranean Sea - Mediterranean CC - bookIRDDocument7 pagesBenedetti 2016 Climate Change Impact On Planktonic Production in The Mediterranean Sea - Mediterranean CC - bookIRDalberte bondeauNo ratings yet
- Frankl CHP10GullydevelopmentintheTigrayHighlandsDocument10 pagesFrankl CHP10GullydevelopmentintheTigrayHighlandsTech TNo ratings yet
- Tomanova EspañolDocument9 pagesTomanova EspañolJose ParraguezNo ratings yet
- Vantrepotte&al2013Document10 pagesVantrepotte&al2013ikramsasa1997No ratings yet
- 2021 - Impacto Dragado en ColumnaDocument11 pages2021 - Impacto Dragado en ColumnaAntonio Bejarano MorenoNo ratings yet
- Danovaro 2018Document17 pagesDanovaro 2018Dominique LefevreNo ratings yet
- 264-Article Text-1004-1-10-20140620 - Fluvial IslandsDocument5 pages264-Article Text-1004-1-10-20140620 - Fluvial IslandsdanielaNo ratings yet
- Seasonal and Spatial Variation in Suspended Matter, Organic CarbonDocument13 pagesSeasonal and Spatial Variation in Suspended Matter, Organic CarbonDr.Abbas HasanNo ratings yet
- Understanding Mn-Nodule Distribution and EvaluationDocument25 pagesUnderstanding Mn-Nodule Distribution and EvaluationwessilissaNo ratings yet
- Identification of The Self-Purification Stretches of The Pinios River, Central GreeceDocument14 pagesIdentification of The Self-Purification Stretches of The Pinios River, Central GreeceSergei OstroumovNo ratings yet
- React Developer with Experience in Web and Mobile AppsDocument1 pageReact Developer with Experience in Web and Mobile AppsJason StanleyNo ratings yet
- Grant Holzemer ResumeDocument1 pageGrant Holzemer ResumeJason StanleyNo ratings yet
- Bro Chemometrics 1997Document9 pagesBro Chemometrics 1997Jason StanleyNo ratings yet
- Sae ApplicationDocument2 pagesSae ApplicationJason StanleyNo ratings yet
- Topic#3.Scalars, Vectors and TensorsDocument64 pagesTopic#3.Scalars, Vectors and TensorsJason StanleyNo ratings yet
- Sejal Divekar Resume1Document2 pagesSejal Divekar Resume1Jason StanleyNo ratings yet
- Efficient Parallel Non-Negative Least Squares On Multi-Core ArchitecturesDocument16 pagesEfficient Parallel Non-Negative Least Squares On Multi-Core ArchitecturesJason StanleyNo ratings yet
- Cublas LibraryDocument254 pagesCublas LibraryJason StanleyNo ratings yet
- Mahesh Bauri Jul2022Document1 pageMahesh Bauri Jul2022Jason StanleyNo ratings yet
- 1 s2.0 S1877050917307858 MainDocument10 pages1 s2.0 S1877050917307858 MainJason StanleyNo ratings yet
- Elijah Hawkins Resume PDFDocument1 pageElijah Hawkins Resume PDFJason StanleyNo ratings yet
- Social Security Numbers For Noncitizens: Does A Noncitizen Need A Social Security Number (SSN) ?Document2 pagesSocial Security Numbers For Noncitizens: Does A Noncitizen Need A Social Security Number (SSN) ?Jason StanleyNo ratings yet
- HW4 PDFDocument23 pagesHW4 PDFJason StanleyNo ratings yet
- PARAFAC. Tutorial and Applications: ElsevierDocument23 pagesPARAFAC. Tutorial and Applications: ElsevierJason StanleyNo ratings yet
- Quadratic Forms and Convexity: Eivind EriksenDocument22 pagesQuadratic Forms and Convexity: Eivind EriksenJason StanleyNo ratings yet
- QR Factorization: Triangular Matrices QR Factorization Gram-Schmidt Algorithm Householder AlgorithmDocument42 pagesQR Factorization: Triangular Matrices QR Factorization Gram-Schmidt Algorithm Householder AlgorithmALEEZA kkNo ratings yet
- Matrix Gradients ExplainedDocument1 pageMatrix Gradients ExplainedJason StanleyNo ratings yet
- LicenseDocument1 pageLicenseRiza IskandarNo ratings yet
- Bounded Variable Least Squares Algorithm Solves Constrained Linear ProblemsDocument13 pagesBounded Variable Least Squares Algorithm Solves Constrained Linear ProblemsJason StanleyNo ratings yet
- OE4020 Assignment - 1: Jason Stanley.S Na15b021Document1 pageOE4020 Assignment - 1: Jason Stanley.S Na15b021Jason StanleyNo ratings yet
- Murray, Knaapen 2008Document18 pagesMurray, Knaapen 2008Jason StanleyNo ratings yet
- B.Tech Curriculum Requirements by DepartmentDocument23 pagesB.Tech Curriculum Requirements by DepartmentBala SudeepNo ratings yet
- Runge KuttaDocument16 pagesRunge KuttaJason StanleyNo ratings yet
- Prediction of Warship Manoeuvring Coefficients Using CFD: C. Oldfield, M. Moradi Larmaei, A. KendrickDocument17 pagesPrediction of Warship Manoeuvring Coefficients Using CFD: C. Oldfield, M. Moradi Larmaei, A. KendrickJason StanleyNo ratings yet
- Fem Shape FunctionDocument11 pagesFem Shape FunctionJason StanleyNo ratings yet
- "List of Possible Topics For OE545 ProjectDocument1 page"List of Possible Topics For OE545 ProjectJason StanleyNo ratings yet
- OE5450: Numerical Techniques in Ocean Hydrodynamics: Implementation of FEM On A Rectangular DomainDocument15 pagesOE5450: Numerical Techniques in Ocean Hydrodynamics: Implementation of FEM On A Rectangular DomainJason StanleyNo ratings yet
- Thermo ScientificDocument29 pagesThermo ScientificTaeng SoshiNo ratings yet
- Impact of Performance Management at InfosysDocument8 pagesImpact of Performance Management at InfosysN Pritam DasNo ratings yet
- Uts 2Document4 pagesUts 2Neill Antonio AbuelNo ratings yet
- (MIÑOZA - Wen Roniel Badayos) - PKYF 2023 Application EssayDocument3 pages(MIÑOZA - Wen Roniel Badayos) - PKYF 2023 Application EssayWen MinozaNo ratings yet
- 3.6.3 Function Block Diagram: 3 Operation TheoryDocument2 pages3.6.3 Function Block Diagram: 3 Operation TheoryAC DCNo ratings yet
- Process data sheet for spent caustic degassing drumDocument4 pagesProcess data sheet for spent caustic degassing drumChakravarthy BharathNo ratings yet
- Fauziah Ratna Hapsari: About MeDocument3 pagesFauziah Ratna Hapsari: About MeFauziah Ratna HapsariNo ratings yet
- SchimmentiDevelopmentalrootsofdissociationpre PrintDocument42 pagesSchimmentiDevelopmentalrootsofdissociationpre PrintZeynep ÖzmeydanNo ratings yet
- How to Write a Resume for a Job ApplicationDocument4 pagesHow to Write a Resume for a Job ApplicationPhuong Dang AnhNo ratings yet
- Listening skills practice: Living online – exercisesDocument2 pagesListening skills practice: Living online – exercisesnavasfernando75100% (1)
- RPT-E3: Analog and Networking SystemsDocument2 pagesRPT-E3: Analog and Networking SystemsenriquealugovNo ratings yet
- Essence of Indian Traditional KnowledgeDocument2 pagesEssence of Indian Traditional KnowledgePavan KocherlakotaNo ratings yet
- Mesozoic Rock Formations of the South-West Rusca Montana BasinDocument27 pagesMesozoic Rock Formations of the South-West Rusca Montana BasinplanessNo ratings yet
- Effect of Halal Awareness, Halal Logo and Attitude On Foreign Consumers' Purchase IntentionDocument19 pagesEffect of Halal Awareness, Halal Logo and Attitude On Foreign Consumers' Purchase IntentionandiahmadabrarNo ratings yet
- Lesson 4 Rainforest CharacteristicsDocument14 pagesLesson 4 Rainforest CharacteristicsLabib Kamal Year 11No ratings yet
- N164.UCPE12B LANmark - OF - UC - 12x - Singlemode - 9 - 125 - OS2 - PE - BlackDocument2 pagesN164.UCPE12B LANmark - OF - UC - 12x - Singlemode - 9 - 125 - OS2 - PE - BlackRaniaTortueNo ratings yet
- CH 2Document28 pagesCH 2CWHNo ratings yet
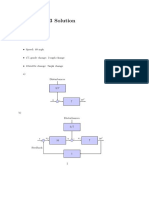
- Homework 3 Solution: Problem 1Document3 pagesHomework 3 Solution: Problem 1Gustavo Felicio PerruciNo ratings yet
- Yousefi 2018Document10 pagesYousefi 2018Yuzar StuffNo ratings yet
- Midsem QP Ce2201 Jan MayDocument2 pagesMidsem QP Ce2201 Jan Maytom meeteiNo ratings yet
- Interesting Facts About AquariusDocument9 pagesInteresting Facts About AquariusFikir FikirNo ratings yet
- 2018 January Proficiency Session 1 A VersionDocument11 pages2018 January Proficiency Session 1 A VersionUğur GiderNo ratings yet
- The Laryngeal Nerve of The Giraffe: Does It Prove Evolution?Document13 pagesThe Laryngeal Nerve of The Giraffe: Does It Prove Evolution?ShwazaraNo ratings yet
- Toxicity Study of Salacia Reticulata Extract PowderDocument9 pagesToxicity Study of Salacia Reticulata Extract PowderAnh LevanNo ratings yet
- Hubungan Penggunaan Gadget Dengan Prestasi Belajar AnakDocument13 pagesHubungan Penggunaan Gadget Dengan Prestasi Belajar AnakIndahNo ratings yet
- 10 Power Phrases To Master Your Next ArgumentDocument6 pages10 Power Phrases To Master Your Next ArgumentshaiannnicolebNo ratings yet
- Siemens Motor PDFDocument177 pagesSiemens Motor PDFณัฐพล นันไชยNo ratings yet
- PNC121 Caring CommunicationDocument32 pagesPNC121 Caring CommunicationMateo Giraldo GomezNo ratings yet