You might also like
- Taxonomy 1349138Document16 pagesTaxonomy 1349138Miguel Angel Perez FarreraNo ratings yet
- Cabrera Et Al. 2008 Araceae PhylogeneticsDocument13 pagesCabrera Et Al. 2008 Araceae PhylogeneticsGerardo A. SalazarNo ratings yet
- Part 281358Document28 pagesPart 281358José ColoradoNo ratings yet
- Carter2014 Sphenoclea Zeylanica (Sphenocleaceae) in North America-Dispersal, Ecology, and MorphologyDocument19 pagesCarter2014 Sphenoclea Zeylanica (Sphenocleaceae) in North America-Dispersal, Ecology, and MorphologyJamesNo ratings yet
- 2011 - Datura Geography - Rev Mex Biodiver - 27851-55123-1-PB PDFDocument12 pages2011 - Datura Geography - Rev Mex Biodiver - 27851-55123-1-PB PDFAlberto VillaNo ratings yet
- Cano Et Al - 2013 - A Revision of Trithrinax PDFDocument53 pagesCano Et Al - 2013 - A Revision of Trithrinax PDFHenoh100% (3)
- Jaltomata y Su TaxonomiaDocument9 pagesJaltomata y Su TaxonomiaLuis GarciaNo ratings yet
- Zamia MexicoDocument36 pagesZamia MexicoPablo GalnNo ratings yet
- 2017 - Yañez Et. AlDocument18 pages2017 - Yañez Et. AlGugú YáñezNo ratings yet
- Analisis Morfometrico de EpidendrumDocument8 pagesAnalisis Morfometrico de EpidendrumPAULA OCAMPONo ratings yet
- Dias Et Al 2015 Metrodorea Phylogeny Bot 1410 49Document10 pagesDias Et Al 2015 Metrodorea Phylogeny Bot 1410 49Fabiana PiokerNo ratings yet
- VazquezLobo AstroDocument10 pagesVazquezLobo AstroTit fuckerNo ratings yet
- Garb 2004Document16 pagesGarb 2004Jonas MartinsNo ratings yet
- Sp. Nov., A New Species in The B. Dothidea-Complex On Eucalyptus in South AfricaDocument10 pagesSp. Nov., A New Species in The B. Dothidea-Complex On Eucalyptus in South AfricamiguelNo ratings yet
- Noben Et Al 2018 Syst Bot Dicksonia NeotropicoDocument20 pagesNoben Et Al 2018 Syst Bot Dicksonia NeotropicoApipa AndinoNo ratings yet
- Silva-y-Eguiarte. Geographic Patterns in The Reproductive Ecology of Agave LechuguillaDocument11 pagesSilva-y-Eguiarte. Geographic Patterns in The Reproductive Ecology of Agave Lechuguillasmricardo2445No ratings yet
- Dracaena L. (Asparagaceae) in The New World: Its History and BotanyDocument23 pagesDracaena L. (Asparagaceae) in The New World: Its History and BotanyMethoz Cuadra RobucaNo ratings yet
- Lankesteriana 9 (3) : 491-504. 2010.Document14 pagesLankesteriana 9 (3) : 491-504. 2010.bahamut58No ratings yet
- Phylogeny and biogeography of Juglans based on molecular dataDocument11 pagesPhylogeny and biogeography of Juglans based on molecular dataDinko BećirspahićNo ratings yet
- Reassessment of Zephyranthes in Yucatán Peninsula including new species Z. orellanaeDocument10 pagesReassessment of Zephyranthes in Yucatán Peninsula including new species Z. orellanaeKaro LinaNo ratings yet
- MaterialesDocument16 pagesMaterialesJason EspitiaNo ratings yet
- Yockteng 2011Document43 pagesYockteng 2011Aline SilvaNo ratings yet
- Trevor J. Hawkeswood- A record of Amanita muscaria (L.) Lam. (Basidiomycetes: Amanitaceae) from Wentworth Falls, New South Wales, Australia with a review of some literature on the ecology of the species within AustraliaDocument3 pagesTrevor J. Hawkeswood- A record of Amanita muscaria (L.) Lam. (Basidiomycetes: Amanitaceae) from Wentworth Falls, New South Wales, Australia with a review of some literature on the ecology of the species within AustraliaGummyColaNo ratings yet
- s10267 007 0364 6Document8 pagess10267 007 0364 6indra maulanaNo ratings yet
- Three New Species and A Nomenclatural Synopsis of Urera (Urticaceae) From Mesoamerica DCJMB4038Document19 pagesThree New Species and A Nomenclatural Synopsis of Urera (Urticaceae) From Mesoamerica DCJMB4038Jorge Hernán Macaya BertiNo ratings yet
- Phylogenyof Dorstenia MoraceaerevealsthepolyphyleticnatureDocument14 pagesPhylogenyof Dorstenia MoraceaerevealsthepolyphyleticnaturePramudito KusumoNo ratings yet
- Marcelo Dias Machado Vianna-Filho Andrea Ferreira Da CostaDocument13 pagesMarcelo Dias Machado Vianna-Filho Andrea Ferreira Da CostaMarcelo ViannaNo ratings yet
- Rydin 2008-Rubiaceae PDFDocument10 pagesRydin 2008-Rubiaceae PDFCarla MaldonadoNo ratings yet
- Phylogeny of Mockingbirds and ThrashersDocument11 pagesPhylogeny of Mockingbirds and ThrashersVermilion~No ratings yet
- Filogenética BasidioDocument2 pagesFilogenética BasidioMartival SantosNo ratings yet
- Phenetic analysis of Mexican pinyon pines reveals species differentiationDocument14 pagesPhenetic analysis of Mexican pinyon pines reveals species differentiationCoraline JonesNo ratings yet
- A Molecular Phylogenetic Study of Graptopetalum CRDocument7 pagesA Molecular Phylogenetic Study of Graptopetalum CRGianna Barcelli FantappieNo ratings yet
- Simon Et Al 2011 American Journal of BotanyDocument21 pagesSimon Et Al 2011 American Journal of BotanyTaline PinheiroNo ratings yet
- 2010 Nototriton Tomamorum TexiguatDocument17 pages2010 Nototriton Tomamorum TexiguatJosue Ramos GaldamezNo ratings yet
- Nova - Hedwigia - 102 - 1 - 2 - 0129 - 0140 - Vitoria - 0293 - WM (MODELO)Document12 pagesNova - Hedwigia - 102 - 1 - 2 - 0129 - 0140 - Vitoria - 0293 - WM (MODELO)Dra. Caroline DamascenoNo ratings yet
- Botanical Society of America, Inc. American Journal of BotanyDocument12 pagesBotanical Society of America, Inc. American Journal of BotanyJulio César Chávez GalarzaNo ratings yet
- Govindarajulu - 2011 Filogenia Diploidia Pob LeucaenaDocument15 pagesGovindarajulu - 2011 Filogenia Diploidia Pob LeucaenaAlma AguilarNo ratings yet
- Soto-Trejo 2015Document15 pagesSoto-Trejo 2015Joel GonzálezNo ratings yet
- Molecular and Morphological Evidence Reveals ThreeDocument25 pagesMolecular and Morphological Evidence Reveals ThreeBruno MarsNo ratings yet
- Fgene 12 742870Document18 pagesFgene 12 742870NicoleNo ratings yet
- Hamamelidaceae AmJBotJuly1999vol86no7 1027-1037Document11 pagesHamamelidaceae AmJBotJuly1999vol86no7 1027-1037geert79No ratings yet
- Botanical Studies .A18Document8 pagesBotanical Studies .A18giangiseasNo ratings yet
- Reproductive Strategies of Venezuelan Palm Swamp PlantsDocument13 pagesReproductive Strategies of Venezuelan Palm Swamp PlantsVinicius da Cruz SousaNo ratings yet
- Guatemalan Apocynaceae family overviewDocument46 pagesGuatemalan Apocynaceae family overviewCristian MéndezNo ratings yet
- El Complejo Sporobolus Indicus (Poaceae, Chloridoideae, Zoysieae) en La ArgentinaDocument12 pagesEl Complejo Sporobolus Indicus (Poaceae, Chloridoideae, Zoysieae) en La ArgentinaGuido BarallaNo ratings yet
- North American Fungi: Puccinia Areolata, P. Treleasiana, and P. GemellaDocument10 pagesNorth American Fungi: Puccinia Areolata, P. Treleasiana, and P. GemellaRicardo SozaNo ratings yet
- Phylogeny of ClematisDocument43 pagesPhylogeny of ClematisSara V. CastañedaNo ratings yet
- Arroyo-Cosultchi Et Al 2006Document12 pagesArroyo-Cosultchi Et Al 2006Reisla OliveiraNo ratings yet
- Diet of The Yellow-Knobbed Curassow in TDocument11 pagesDiet of The Yellow-Knobbed Curassow in TDiegoNo ratings yet
- Pdis 92 4 0519Document11 pagesPdis 92 4 0519Samuel Paucar CamposNo ratings yet
- Taxon - 1979 - 28 - Rzedowski PDFDocument16 pagesTaxon - 1979 - 28 - Rzedowski PDFWilliams Marcel Caceres FerreiraNo ratings yet
- Arias Et Al-2010Document12 pagesArias Et Al-2010Rpfnm EtnobiologíaNo ratings yet
- Conzattii Standley and D. Riojae From Across Its Known Range of Cultivation. Diospyros Riojae IsDocument24 pagesConzattii Standley and D. Riojae From Across Its Known Range of Cultivation. Diospyros Riojae IsRoyer Garra de OsoNo ratings yet
- Craugastor Rhodopis C-Occidentalis Group Philogenetics Streicher Etal 2014Document22 pagesCraugastor Rhodopis C-Occidentalis Group Philogenetics Streicher Etal 2014Paulino PonceNo ratings yet
- MPE Amazonian Dactyloa AnolesDocument11 pagesMPE Amazonian Dactyloa AnolesRicardo VasquezNo ratings yet
- Biogeographic Analysis of Endemic Cacti of The SieDocument18 pagesBiogeographic Analysis of Endemic Cacti of The SieDavid BrailovskyNo ratings yet
- H ViridssimumDocument23 pagesH ViridssimumReyna GutiérrezNo ratings yet
- A Revision of Erato Compositae LiabeaeDocument14 pagesA Revision of Erato Compositae LiabeaeLuis Garcia ChisculNo ratings yet
- Phylogeny of The Neotropical Tribe Jacarandeae (Bignoniaceae)Document13 pagesPhylogeny of The Neotropical Tribe Jacarandeae (Bignoniaceae)Francione GomesNo ratings yet
- Piperno and Flannery 2001 PDFDocument3 pagesPiperno and Flannery 2001 PDFAlberto VillaNo ratings yet
- Delgadoet Al. 2003 - MesoamericaDocument13 pagesDelgadoet Al. 2003 - MesoamericaAlberto VillaNo ratings yet
- Everything You Need to Know About Edible Wild Greens Used by Indigenous GroupsDocument15 pagesEverything You Need to Know About Edible Wild Greens Used by Indigenous GroupsAlberto VillaNo ratings yet
- 2013 - Molecular Phylogeny Datura - Syst BotDocument12 pages2013 - Molecular Phylogeny Datura - Syst BotAlberto VillaNo ratings yet
- 2018 Benítez Et Al. The Genus Datura L. (Solanaceae) in Mexico and Spain PDFDocument19 pages2018 Benítez Et Al. The Genus Datura L. (Solanaceae) in Mexico and Spain PDFAlberto VillaNo ratings yet
- Plant Standard CFIADocument84 pagesPlant Standard CFIAGhassan LafiNo ratings yet
- Cell CycleDocument20 pagesCell CycleShelrenNo ratings yet
- Biological Basis of BehaviorDocument27 pagesBiological Basis of Behaviorhafizamanzoor44No ratings yet
- Senarai Jurnal Malaysia Dalam Pangkalan DataDocument22 pagesSenarai Jurnal Malaysia Dalam Pangkalan DataNani Herlin JohannNo ratings yet
- Form Ii Biology End Term of Term IiDocument7 pagesForm Ii Biology End Term of Term IioyooNo ratings yet
- WEINER Aesthetics Is A Cross Cultural CategoryDocument38 pagesWEINER Aesthetics Is A Cross Cultural CategoryjoseanapNo ratings yet
- National Park ThesisDocument4 pagesNational Park Thesisjuliepottsdenton100% (2)
- General Biology 2 - Grade 12Document16 pagesGeneral Biology 2 - Grade 12Candice LopezNo ratings yet
- Genitourinary SystemDocument25 pagesGenitourinary SystemAlen Vukosavljevic100% (1)
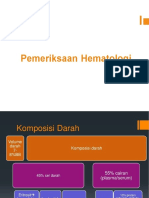
- Pemeriksaan Hematologi PDFDocument47 pagesPemeriksaan Hematologi PDFilham perdiansyahNo ratings yet
- DNA Replication SlideshowDocument14 pagesDNA Replication SlideshowRennie SmolenskiNo ratings yet
- Table of Specifications: 2Nd Quarter ExaminationDocument2 pagesTable of Specifications: 2Nd Quarter ExaminationLeah BobisNo ratings yet
- Review of Literature 2.1. Online Resources For The Plant TaxonomyDocument7 pagesReview of Literature 2.1. Online Resources For The Plant TaxonomyRhoda Mae CubillaNo ratings yet
- CV StefLhermitteDocument16 pagesCV StefLhermitteAgronomistapoloNo ratings yet
- Daraga Community College: Commission On Higher EducationDocument2 pagesDaraga Community College: Commission On Higher EducationJodelen B. Percol0% (1)
- LSM1102 Cyh Lecture 2 FaqDocument5 pagesLSM1102 Cyh Lecture 2 FaqtxNo ratings yet
- Cambridge IGCSE: BIOLOGY 0610/42Document20 pagesCambridge IGCSE: BIOLOGY 0610/42mahmoud shaheen100% (1)
- Moonwalk National High School: Schools Division of Parañaque CityDocument4 pagesMoonwalk National High School: Schools Division of Parañaque CityRowena TabisoraNo ratings yet
- Evolution, Science, and Society: Evolutionary Biology and The National Research AgendaDocument75 pagesEvolution, Science, and Society: Evolutionary Biology and The National Research Agendaজুম্মা খানNo ratings yet
- Cell Growth, Division, and Reproduction: Limits To Cell SizeDocument2 pagesCell Growth, Division, and Reproduction: Limits To Cell SizeZnite100% (1)
- Cell Modifications Help Cells FunctionDocument2 pagesCell Modifications Help Cells FunctionJasmine FloreceNo ratings yet
- Iloilo Science and Technology University Grade 3 Science SpecificationsDocument6 pagesIloilo Science and Technology University Grade 3 Science SpecificationsMicayNo ratings yet
- Nucleotide Sugar Transporters in Golgi ApparatusDocument8 pagesNucleotide Sugar Transporters in Golgi ApparatusCosmin BarbosNo ratings yet
- CamBeads Si Usage InstructionsDocument5 pagesCamBeads Si Usage Instructionsrohit asilNo ratings yet
- Extra Questions For Class 9th - CH 5 The Fundamental Unit of Life Science Study Rankers PDFDocument4 pagesExtra Questions For Class 9th - CH 5 The Fundamental Unit of Life Science Study Rankers PDFsssNo ratings yet
- Biotech 3 Cell DivisionDocument4 pagesBiotech 3 Cell DivisionClinton YmbongNo ratings yet
- Forensic 1Document3 pagesForensic 1Happy Jane YanguasNo ratings yet
- List of Advantages and Disadvantages of NanotechnologyDocument4 pagesList of Advantages and Disadvantages of Nanotechnologyachilles. chixxNo ratings yet
- Republic of The Philippines: Tuburan CampusDocument3 pagesRepublic of The Philippines: Tuburan CampusFaith MaratasNo ratings yet
- Annamalai University: Paper - I: Fundamentals of Sociology - I Code No. D - 56Document147 pagesAnnamalai University: Paper - I: Fundamentals of Sociology - I Code No. D - 56Joseph JohnNo ratings yet
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet